
迈克尔·贝希关于鞭毛的观点并未被推翻
乔纳森•麦克莱奇(Jonathan McLatchie) 2011年3月15日 翻译:张跃江
原文见 Michael Behe Hasn’t Been Refuted on the Flagellum | Evolution News

迈克尔·贝希(Michael Behe)曾经就细菌的鞭毛提出过“不可化约之复杂性”的论证,而进化论者也给出了标准的达尔文式的反驳。我们中间凡是对智慧设计与进化论之争有所涉猎的人,对此想必耳熟能详。
在进化论理论家中似乎存在一种共识,即细菌鞭毛不可化约之复杂性已经被推翻了,而我们这些智慧设计者只是在无谓地不断转移目标,把头埋在沙土之中,企图抓住最后的救命稻草。
事实上,有人最近在社交媒体“脸书”上这样评论:
“我对智慧设计的信奉者们最不耐烦的就是他们似乎永不承认失败。多少次你们被告知这是错的,你们才接受呢?智慧设计论在同行评议的刊物上被驳斥多少回,你们才会放弃这个已经失败的主张?细菌鞭毛不可化约的复杂性故事已经完全而彻底地死掉了。翻篇吧。”
最近,我在一场科学与信仰交流会的午餐问答环节中,提及细菌鞭毛不可化约的复杂性的时候,遭遇了类似的回应。
然而,这种宣称是真的吗?贝希论证真的被批评家们推翻了吗?大约一年前,我读了一本书——《为什么智慧设计论失败了—对新创造论者的科学批判》(Why Intelligent Design Fails – A Scientific Critique of the New Creationism, 编者Matt Young和Taner Edis)。该书第五章“细菌鞭毛的进化”由伊恩·马斯格拉夫(Ian Musgrave)撰写。作为针对迈克尔·贝希和威廉·但布斯基(William Dembski)的回应,马斯格拉夫试图彻底终结不可化约之复杂性的主张。读完他所写的一章,我认为了无新意。在书中第82页,马斯格拉夫为我们提供了下列论述:
“真细菌鞭毛进化的可能图景是这样的:一种基于SMC棒和成孔复合体的分泌系统首先出现,这是III型分泌系统与鞭毛系统的共同祖先。离子泵(后来变成了马达蛋白)与该结构的结合提升了分泌功能。直至今日,马达蛋白作为分泌驱动蛋白家族的一员,仍能自如地与鞭毛结构分离或者重新结合。在这个阶段,棒和成孔复合体可能已经能够旋转,就像某些滑动系统中的棒状结构一样。下一步出现的是原始鞭毛丝,成为蛋白分泌结构的一部分(试比较假单胞菌的菌毛、沙门氏菌的和大肠杆菌的丝状结构)。到这一阶段(或者再晚一点)就能够滑动或扭动,经过改进就可以游动。调节与切换可以后来再加上,因为有些现代真细菌缺乏这些性能,但也能适应其生存环境(Shah and Sockett 1995)。在每个阶段,结构上的变化总能带来一项益处。”
事实上,马克•帕乐(Mark Pallen)和尼克•马兹可(Nick Matzke)2006年在《自然综述》上发表的文章里已经有类似的论述(最近贝希访问英国时一位听众提及这篇文章)。而且,肯•米勒(Ken Miller)也因为经常做出类似的论断而成名,他主要是基于蛋白质序列的同源性而宣称鞭毛是从III型分泌系统进化而来。
那么,这些论点是否成功地彻底终结了让他们烦脑的智慧设计论?实际上没有;他们没有做到。事实上,我得指出,以上提及的先生们所给出的论证从根本上淡化了几个重大问题。
第一点,也是最重要的,他们淡化了鞭毛系统的高度复杂性和精巧程度——无论是其组装系统还是其前卫的结构设计。事实上,细菌鞭毛在细胞内的自我组装过程是如此地复杂,以至于我在很长时间内都纠结于如何才能向外行人讲明白。其中包含的核心概念,对于不熟悉这个系统或者第一次接触的人来说,是骇人的难懂。另一方面,鞭毛自我组装的机械基础又精巧得令人屏息、令人痴迷;仅仅是鞭毛动力部件绝妙的工程设计就为达尔文主义带来了巨大的挑战,这一点需要对鞭毛马达的运作方式有起码的知识才能正确地理解。让我们来一窥究竟。
鞭毛复合体的自我组装
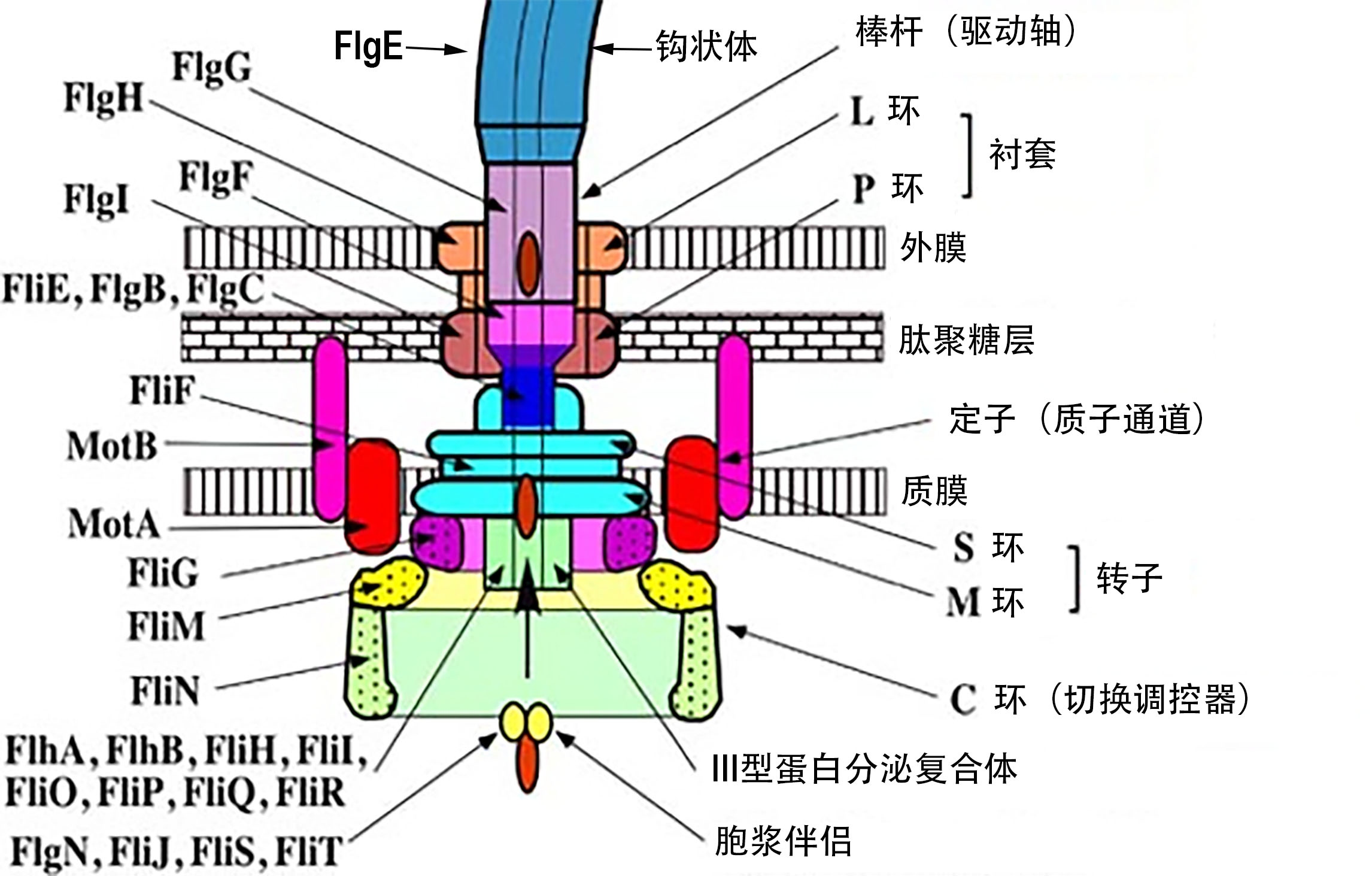
细菌鞭毛的合成需要60多个基因产物的协调表达。它在细胞内的生物合成受控于一套严密组织的基因,环环相扣,层次分明,每个层级上的基因表达都依赖于上一级基因的率先表达。鞭毛组装的典范物种是肠杆菌科的沙门氏菌。下面的论述如果不特别指出,都主要是指沙门氏菌。
沙门氏菌鞭毛系统有三类启动子(启动子类似于一种分子开关,当启动子被RNA聚合酶和与之相关的被称为σ因子的特殊蛋白质识别时,就能够启动基因表达)。 这三类启动子被简单地命名为第一类、第二类和第三类。三类启动子控制了三组基因的依次表达,在顺序上与鞭毛组装的过程紧密关联。第一类只控制一个操纵子内的两个基因,FlhD和FlhC。第二类控制8个操纵子内的35个基因,包括与钩状体-基体、其他鞭毛部件和输出复合体的组装相关的基因,以及两个名为FliA和FlgM的调节基因。与丝状体合成相关的基因则由第三类启动子所控制。
第一类启动子驱动一个称为FldH4C2(记不住它也不要担心!)的主要调节因子的表达(见于包含沙门式菌属的肠杆菌科)。然后,该调节因子与一个σ因子(σ70)结合,打开第二类启动子(如上所述,σ因子是一种蛋白质,能使RNA聚合酶与特定基因的启动子结合)。接着,第二类启动子负责钩状体-基体亚单位及其调节蛋白的表达,这些调节蛋白中包括了另外一个σ因子, 即σ28(其基因称为FliA)及其反σ因子,FlgM (反σ因子,顾名思义,能与σ因子结合并抑制其转录活性)。σ28是激活第三类启动子所必需。但这样我们陷入一个潜在的困境。在完成钩状体-基体的建造之前就开始表达鞭毛蛋白单体是绝对没有道理的。为了抑制σ28, 上述反σ因子(FlgM)会压抑σ28的活性,禁止它与RNA聚合酶复合体发生作用。在钩状体-基体构建完毕之后,由第二类基因所编码的鞭毛结构将反σ因子FlgM分泌到细胞外。第三类启动子(负责表达鞭毛单体、趋化系统和马达动力机关)才终于被σ28激活,从而完成鞭毛的建造。
还有比这更妙的。鞭毛输出系统(FlgM被移出细胞的通道)有两种底物特异性状态:棒杆/钩状体类底物和丝状体类底物。在鞭毛组装过程中,底物特异性开关需要从前一种状态切换到后一种。构建钩状体和棒杆的蛋白质部件的输出要先于丝状体部件。但这种底物特异性的转换是如何发生的呢?
一种叫做FlhB的膜结合蛋白在此过程中发挥关键作用。还有一种名叫FliK的鞭毛钩长度调节蛋白,它负责将钩状体的长度控制在55纳米左右。这种蛋白质同时负责切换运输底物的特异性。结果呢,没有了FliK,就不能切换底物以输出丝状体,也失去了控制钩状体长度的能力。FliK有两个关键的结构域, 即N-末端(FliKN)和C末端(FliKC)。在钩状体组装过程中,FliKN充当分子感受器和长度信息传送器。当钩状体达到恰当长度的时候,信息被传递给FliKC (具体地是一个称为FliKCT的亚结构),导致FliKC构象变化,致使FliKC与FlhBC结合。这又导致FlhBC(FlhB的一个结构域)发生构象改变,从而造成底物特异性的切换。

鞭毛组装开始于细胞质膜,经过壁膜间隙,最终延申至细胞外。鞭毛基本上包含两个主要部分:分泌系统和轴向结构。轴向结构的主要部件是FlgG棒杆、FlgE钩状体和FliC丝状体。所有这些部件的组装都需要一种帽蛋白(分别是FlgJ、FlgD和FliD)的协助。其中,只有FliD会在最终产品中保留于丝状体的顶端。其他轴向部件(被称为FlgB、FlgC和FlgF)将棒杆和MS环形复合体相连接。钩状体和丝状体由FlgK和FlgL连接在一起。
当C环与C杆在细胞质膜表面与M环粘结之后,整个鞭毛复合体的结构基础——MS环形复合体——就建成了,它开始向外分泌鞭毛蛋白。
棒杆结构是穿越肽聚糖层而建造的,但它的延申要冲破外膜的物理屏障,还是需要特别的帮助。于是,外环复合体在膜上切出一个洞,以便钩状体能够在FlgD脚手架内长出,直到长成55纳米的长度阈值。然后,MS复合体向外分泌的底物从棒杆-钩状体模式切换成鞭毛蛋白模式,FlgD可以被钩状体附属蛋白所取代,这样丝状物就可以持续生长。如果没有帽蛋白FliD, 鞭毛蛋白单体就会丢失。所以帽蛋白是这个过程所必须的。
为什么III型分泌系统不会进化成鞭毛
有人也许认为以上描述已经足以证明肯•米勒等人大手一挥的说法是极端的简单化了,但是达尔文主义故事中的问题还不止于此。为什么鞭毛生物合成过程是如此精准地调节和紧密协调的呢?不仅能量需求使得鞭毛系统的运行极为昂贵,而且鞭毛蛋白不合时宜的表达还会引发宿主系统强烈的免疫反应,细菌可不乐意这样做。
从进化的思路来看,这有什么意义?而且鞭毛蛋白单体还能强力地诱生细胞因子。假设你是一个拥有III型分泌系统的耶尔辛氏菌,你最不希望做的就是把鞭毛蛋白的肽链呈现给巨噬细胞。毫无疑问,这会损害耶尔辛氏菌的抗炎机制。
结论
我前面的描述仅仅触及了鞭毛这个壮观的纳米技术产品的皮毛。为了简洁,我甚至没有讨论高超的趋化过程、双部件信号传递环路、旋转方向切换和为鞭毛提供能量的质子动势。归根到底,现代达尔文理论,根据其经典理解,无论如何无法解释如此高超的复合体及其精致的动力装置是如何起源的。就像对于眼睛起源的达尔文式“解释”,对于没有学习过、对视觉的生化和分子水平上的工程奇迹一无所知的人,乍一看似乎有道理。同样地,当考虑了鞭毛系统的分子细节之后,进化论的“解释”就很快变得苍白无力。当你通盘了解了上述细节,以及新达尔文主义无力解释新颖的蛋白质折叠和蛋白质-蛋白质结合位点之后,你真的会相信整个系统可以经由细微的、渐进的改变,一次一小步地拼凑出来吗?按着新进化论主义的主要卖点,它声称有能力消除处处可见的设计痕迹,难道不能根据它明显的无力再次让智慧设计论成为可供选择的、值得尊重的科学提议?
生物研究所的道格拉斯•阿克斯(Douglas Axe)最近发表在《生物复杂性》期刊(Bio-complexity )撰文指出,基因复制和旧材新用只在不需要多少改变就能获得可供选择的新功能的情况下才有可行性。如果基因复制是中性的(对生物体的代价),那么在细菌群体中产生新功能所需要的突变不可以超过六个。如果复制的基因对适应能力有一丝负面影响,所允许的最大变异数目就降到2甚至更低(不包含基因复制本身)。
今天细菌鞭毛对于达尔文主义的挑战,比贝希1996年初次出版《达尔文的黑箱》的时候并没有减弱,甚至更大了。
作者简介
乔纳森•麦克莱奇博士拥有斯特拉斯克莱德大学(University of Strathclyde)司法生物学学士、格拉斯哥大学(University of Glasgow)进化生物学硕士、新堡大学(Newcastle University)医学与分子生物学硕士和进化生物学博士学位。目前,他就职于马赛诸塞州波士顿的萨特勒学院(Sattler College),担任生物学助理教授。乔纳森博士曾多次接受数家播客和电台专访,比如首要基督徒电台(Premier Christian Radio)的“难以置信?”节目等。他还在欧洲、北美、南非和亚洲演说,倡导大自然中存在智慧设计的证据。
