
原文网址
https://creation.com/dinosaur-blood-fuz-rana
作者:马克·阿米蒂奇Mark Armitage 翻译:Joe Jack
从蒙大拿州地狱溪( Hell Creek)的三角龙之角里发现的纤维骨软层以及拥有惊人保存度的骨细胞,无法用法扎勒·拉纳(Fazale Rana)博士在其《恐龙血液与地球年龄》一书中提出的成因来解释。本篇将检视并纠正拉纳对三角龙角里软组织的错误认识和错误定性。仅鹦鹉学舌般地重复进化论者对恐龙软细胞的讲法并不能解释这些东西的存在。所以需要进行更为具体的研究工作才能解释之。
拉纳所著《恐龙血液与地球年龄》[1]一书的本质,就是拉纳博士在重复一些世俗科学家对于保存完好到令人咂舌的恐龙软组织的见解,而这些“解释”却诉诸于一些此前未知的“机制”。因此只要这些所谓的机制被否定了,就理当让世俗学界对此闭嘴,且同时也把解释的压力转移到了他们那边——是他们才需要再去做更具体的工作,以期解释这些细胞何以健在。
这些属于生物本身的内源性分子、蛋白质、纤维骨片层、完好的血管、以及不可思议保存度的恐龙骨细胞(osteocytes)等,对拉纳博士及其信徒可是个大问题。毕竟,一般人的生活经验也直觉地告诉他们这些东西与漫长历史相矛盾:既然恐龙骨头是“如此地古老”,它们在这亿万岁月里定然历经了太多的侵蚀腐蚀、食腐生物、以及其它破坏过程,以至于一般人“知道”这不可能。
拉纳必须确保他的信徒对于恐龙软组织拥有处惊不乱的态度。他与休·罗斯(Hugh Ross)的信仰侍奉工作乃奠基于反科学反圣经的假设上,即认为地球已经有数十亿年历史了。因此现在他必须向其信徒解释,为何从亘古的动物身上提取的软组织一点都不稀奇。
似乎是为了淡化这显而易见的事实,拉纳写道,“科学界没多少人对这种论点感到有什么大不了(在其书引言11页)——即软体组织的保存表明了这些生物遗体乃是年轻的。”——然而我可以肯定地告诉你,在加州州立大学(California State University)的生物学家们对此感到了心理冲击。因为能在开裂且含水的三角龙角里发现软组织,是对漫长时间范式的打击。
我对加利福尼亚州的法律起诉,可以证明就在我发表的三角龙角的软组织论文上线的当天,几个有权势的生物系教授便互相接头并进而决定终止我所主管的百万美元级显微学实验室的本人职务。况且专门研究恐龙软体组织的科学家本就人数不多,这主要是由于这可能会毁了他们的职业生涯。由此可见与拉纳的说法完全相反,很多科学界的人士了解恐龙软细胞与软组织的存在所能引发的明显后果。
拉纳声称他此书的目的是为了帮助基督徒从一个“生化学家的角度”(12页)明白,为何软组织残骸能在被断定为亿万年的化石中保存(12页)。他称自己了解“分子的结构、功能、与稳定性。”,并感觉自己有资格去防止意愿良好(但可能是无知)的信徒为“年轻地球论去做有科学问题的论证”(12页)。
拉纳也写道他此书的第二个目的是帮助基督徒“克服信仰年老地球创造论的障碍”(13页)——而这实际上才是此书的真正目的,因为现在从所谓的远古生物残骸里发现软体组织几乎已经是普遍且日常的事了。
遗憾的是这里发文有字数限制,故而我对拉纳的书只提供一个缩减的评述。读者可以发电子邮件(micromark@juno.com)索要完整版的评述。另外也有研究可以表明拉纳对于经文的立场也是错误的。[2]
说这本书肯定是一本反对年轻地球创造论的书其实还是说轻了。拉纳在书里时不时光火并贬低年轻地球创造论,而这就表明我们已经成了他试图约束信徒不越线、不去质疑他关于年老地球论教导的最大障碍。实际上这本书的引言、每一章、结论、乃至附录都有特别针对我们所持的年轻地球论。
在引言大约到了第五段,拉纳开始攻击起了年轻地球论者。一开始他就把年轻地球论者与他所谓的真科学对立起来,并称年轻地球论在他所谓的真正科学事实面前只有一点点道理。
在拉纳开始攻击年轻地球论的开场白之后,我旋即便成为被他点名的第一个创造论科学家——这并不奇怪,因为至少这点拉纳是对的:他如实报道了是我发表了关于软组织的原创科学发现,我也是第一个报告了三角龙角里的软组织。[3]不过,你们从我接下去对第4章的评述里会看到,拉纳暴露了他对于这些研究工作之真正意义的无知。
一个惹眼的暴露拉纳无知的例子就是他用了“软组织残留”这样的称呼(光引言中就出现了11次)。因为这就展现出他对于令人震惊的、数量极多的、保存度不可思议的骨细胞的明显无视——而我不仅在三角龙里,也在矮暴龙(Nanotyrannus)椎骨和跖骨(图1、2)中找到了这些。(矮暴龙最近被定为一个与霸王龙的类似的属)
放射性测年
该书的第三章回顾了放射性定年法。在这个议题上,早已经有了很多优秀的从年轻地球论及圣经观点出发的研究资料对此进行了探讨以及揭露。[4]不过恐龙软细胞的惊人保存状况看来已经“击溃”了这些放射性测年法,因此拉纳便不得不在这个环节去捍卫该方法。以至于他甚至在12页写道,“第三章的目的是为了证明为何放射性检测法‘确实’可靠(强调号在其他原文中就有)。“——而假如放射性检测法真可靠的话,他再做这种陈述称就显得颇为别扭尴尬。
不过拉纳依然一头扎进其中,在第三章中大费周章、彻底评述了放射性定年法的复杂性和技术性,并提到了该方法需要几年工作技能的专家(3章、44页)以及对此法充分了解的地球化学家才能理解并做对(45页)。然而对于此检测法可具有“误导性”并“偶尔确实存在可改变放射衰变的化学和物理过程”(第41页),拉纳却只是一笔带过。很遗憾地是,拉纳选择不把已经被观察到的太阳可影响一些核衰变速率的事实[5], [6]与他读者分享。如果我们现在已能发现诸如太阳因素等能影响一些衰变速率,就保不定尚有未知的其它情况也能改变这些“恒定”的速率。
而正是在这一章,我必须要纠正雷纳的两点关键错误。第一点是他明显的认识错误,他说“阿米蒂奇是用温和的酸浴浸泡了部分三角龙的龙角化石的一个月之后,才提取了柔软、灵动的大约20厘米长10厘米宽的棕色软层。”(3章50页)
然而三角龙角可没有变成化石(没有矿化)。[7] 它可以对病理学实验室常用来脱钙的弱酸性乙二胺四乙酸(EDTA)产生反应且发生脱钙(EDTA其实是用来除去骨骼磷灰质晶格里的钙)。再者,拉纳仅需要读过论文摘要里第二句话,就该明白他关于把骨头浸泡在酸里以及“柔软、灵动的棕色片层”的描述更是错到了离谱。或者他也仅需阅读了我论文图5的文字描述也行,即,“光镜显微照片:从‘未脱钙的三角龙’骨头样本(黑色箭头)中剥离的软组织皮瓣(白色箭头)”。
而就算这些他都没看到,那么我论文在604页描述到:“大块长条状的细轻、浅棕色柔软物质(20厘米长10厘米宽),是从其它固定或未经固定的未脱钙的角骨部分的最里面所提取(粗体是我加的)。” 这即是说,拉纳所指的“软的、灵动的棕色片层”乃是在未被浸泡的骨头里被发现的——提取这些软体组织根本没必要把这些骨头浸泡在任何东西里,它们就靠着硬骨呆在那里。
第二点,拉纳想当然地以为含有“软的、灵动的棕色片层”的角接受了碳14检测,其结果由托马斯(Thomas)与尼尔森(Nelson)发布。[8]
然而这不是事实。我知道哪些送检了碳14,哪些没有。拉纳在这个错误的基础上继续对创造论推论道,“所检测到的碳14不可能是原生性的(内生性的)”。他争辩道,“如此多的又软又灵动的棕色物质理当产生强烈的碳14信号”,并且“报告的数据与任何将灵动的软组织片层视为只有3000至6000年历史的说法无法协调一致”。(50页)
可是送检碳14的三角龙角部分(在最初的碳14检查中)并不是在其中发现了柔软且灵动的棕色片层的纤维骨的中心部分。我所送检的其实是经历各种侵蚀、开裂的25厘米直径的角的外层部分。至于检测结果比预期更古老并非意料之外,因为送检的样本在其埋葬的土里是上下颠倒的姿态,其中的血管成分暴露于降雨、植物根系、微生物、昆虫等等的作用下——这些都会降解外层骨头的软组织。
而我尚没有把软层送检碳14。所以拉纳在写书时其实对究竟送检了什么根本不知道,但他至少应该问一下啊。因此,他关于应该有更强的碳14信号,但检测却得出比预测的碳14年龄更古老的又软又灵活的棕色纤维骨片层的论证,完全不成立。(50到51页)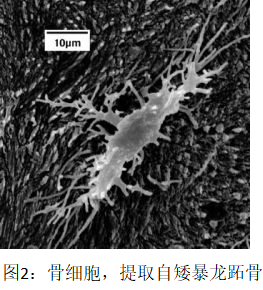
此外,拉纳也忽略了证据确凿的学术文献中关于在所谓数百万到数十亿年历史的贝类化石、骨头、土壤、煤炭、以及钻石中也发现了碳14的强信号。[9] 而拉纳在他书里的第4章,特别是关于2013年《组织化学学报》(Acta Histochemica)论文里讨论的骨头软纤维片层的部分,更是一大败笔。拉纳争论道:“在恐龙化石中保存的大部分生物分子可归因于两个性质:1,广泛的交叉连接(cross-linking) 2,与石墨相似的化学构成。
拉纳关于胶原蛋白具有大量交叉连接的说法虽然并非不正确,可用石墨来做类比就颇为误导一些民众了。许多生物组织的特点就是众多重复多聚体亚基的连接。由于石墨的化学键远比生物组织牢固,因此石墨被认为具有更为复杂的结构,但它本身并非生物组织。更关键的是骨细胞可不是胶原,它们不仅脆弱且其结构中不存在交叉连接。同样的情况也适用于DNA,它们也不存在什么交叉连接。然而拉纳可不顾这一切,依然用7种所谓“牢固”的生化结构去尝试解释在浅墓中的恐龙软组织可历经亿万年的暴露而照旧存在。
我注意到拉纳在这里几乎没有谈论到水分子以及氧化剂对曾经活着的生物组织系统的高度破坏作用(例如游离铁所产生的破坏作用)。更重要的是拉纳所谈到的7种牢固的化学结构压根与细胞膜没有关系。我从恐龙尸骸中所发现的那数千个骨细胞都有细胞膜,而每个骨细胞的磷脂双层膜都极易被水和氧化剂严重破坏降解。[10]
可令人感到奇怪的是拉纳颇为努力地去解释血红素分子的坚固性,并总结道,“ (将铁原子固定在中心的)卟啉环是极为稳定的化合物,这有助于解释它们在恐龙化石中的存在。”(第61页)
只是拉纳这样做反成了对施怀哲博士(Dr.Schweitzer)所提出假设的极好驳斥,即游离的铁原子可通过芬顿化学反应而产生氢氧自由基和过氧化物(氧化剂),进而以某种方式像甲醛一样“固定”了软组织。
可假如血红素分子真如此稳定,里头的铁又怎么可能被释放出来?此外,生物组织又如何能在不被危险的氢氧氧化剂破坏之前,为之“固定”?所有这些施怀哲团队[11]和拉纳都没有解释。[12]
在第62页,拉纳承认目前他只有半吊子的解释,因为“稳定性本身还不足以解释亿万年化石里的软体组织”。然后他又强调了这些话的重要性:“必须同时还要满足其它许多条件。”——可他列举的9个必要的稳定条件没有一个与我们获取的三角龙角有任何关系。
必要的稳定条件
现在让我们考察9种“稳定条件”
条件1
“在石化过程中(我认为拉纳这里指的是矿化过程),富含矿物的水会渗透生物遗骸……而骨头和其它部分里的原生矿物质会被来自外在环境中的矿物所替换”(62页)。在这三角龙角的案例中,只有暴露于外在环境(其中包括富含矿物的水)中的血管才硬化为石头。这就是为什么它们对脱钙法没有反应。然而这些骨头却依然是骨头。这点可在我论文的图14与图15中清楚看到。[13] 用国内任何一家病理实验室所采用的检查人类骨骼中软组织的规范操作去脱钙,这些骨头都会有反应。因此这些骨头依然还是骨头。故而此条件无效。
条件2
“埋藏的条件似乎也有其作用。设想中,水会更容易从生物遗体中流走,由此创造的更干燥环境便除去了微生物和环境酶”(62页)。可在龙角的情况下,我的论文清楚地论述道:“当角被获取时并非处于干燥状态,且其实还有泥泞的基质深深嵌于其中”(p.66)。埋葬地确实有更干燥的条件,但角可不是这样,我们在论文中几处描述了角是湿的。因此它当然充满了细菌、微生物、以及环境酶。由此可见,此条件二无效。
条件3
就第三个条件,拉纳探讨了“干燥、无水条件”,这点我们上文已经谈过了。不过此处他似乎又开始论证起潮湿样本的必要性了!“有趣的是,在一些事例中,有限量的水或许真有助于胶原等生物分子的保存”(63页)。那我倒要问了,到底是湿的还是干的?拉纳对此的回答却显然是——亦湿亦干!
条件4
就这个条件,拉纳把氧视为“极为活跃、具有破坏性的化学作用、且随时随地破坏有机体”(64页),因此软组织必须在氧被隔绝的情况下才能保存下来。而在我《组织化学学报》(Acta Histocheca)上的论文的几处(包括插图),7我们观察到了(甚至在一些柔软棕色软层组织的底部也)布满了植物的根系,这或许也促成角的开裂。因此,氧可以达及开裂骨头的深处。然而大片纤维骨、及保存精致的骨细胞却依然存在。因此条件4无效。
条件5
关于第五个稳定条件,拉纳强调了软组织残骸必须远离环境因素,否则例如消化酶及其它化学物质就会摧毁它们。常用的一个论证是软组织由于被硬骨包裹而得到保护。因此起破坏作用的酶无法触及软组织。然而程序化的细胞死亡以及单纯的熵效应就会让缺乏供养与照料的细胞自行腐烂,这无关乎它们是否嵌在骨矿物质里。因而它们当初只有迅速地被保护起来,我们才能在今天观察到保存度让人震惊的细胞。由此,要让“铁保存”理论行得通且快速发挥作用,细胞与生物组织就必须能够接触到游离铁,并在水的参与下,藉由芬顿化学反应条件下所产生的本会破坏软组织的氢氧自由基与过氧化物再去稳定这些软组织。如果是这样,那么一定发生了奇迹,以某种方式让我们今天能观察到的软组织在隔绝于酶解作用及水解作用的同时,又利用这些化学物质发挥“稳定”作用。而“游离铁”必须从血红素中剥离出来。可血红素却是“极为牢固的化合物”(61页),这又反过来阻止了铁的“游离化”。这“游离”的铁还必须在有水的条件下避开氧,否则便会形成氧化铁化合物。更有甚者,我们还另外再需要一个奇迹:即这一切必须要在20分钟内完成,否则血液一旦凝固就没啥游离铁了。拉纳也观察到了微生物的存在,而这再次敲响了软组织能够长久保存的丧钟。并且又一次,血红蛋白据称又要奇迹般地充当多个角色,包括抗菌、多功能消毒剂、保护剂、稳定剂与甲醛固定剂……方能在解决了需解决的问题的同时还回避了需回避的问题。如果真有这样的事,那就等于你既吃掉了软组织又要想保留了软组织。
条件6
正是在这个环节,拉纳完全认同起适才讨论的“铁保存”实验。就这个实验而言,几乎每个创造论科学家都视为臭名昭著且过于天真;而到了进化论者那里,引用该实验则是用来噤声那些认为亿万年之久的惊人细胞与软组织不可能宛如神迹般被保护下来的反对言论。如果像拉纳这样的进化论者要维护科学界的信誉,他就必须回答我对于该实验的一些反对理由。
6.1
施怀哲等人的“铁保存”实验论文11里的材料与方法算是最说明问题了(有一段时间,这一部分只能在搜索《皇家学会会报B辑》[Proceedings of the Royal Society]网页时才能看到)。然而其中所牵涉到的材料准备以及试验方法“电子版补充材料”,至少根据施怀哲等人论文里提供的链接,却已经无法在网上查阅了。尽管如此,我们依旧能发现她们为了获得“游离”铁而付出的各种人为努力是极为繁复的。首先,鸡与鸵鸟的血液必须与乙二胺四乙酸(EDTA)相混合,以防止血液凝固。然后血液又要被放到高速离心机上进行了多次离心,才完全移除血浆(其中含有凝血蛋白和酶)、血小板及所有白细胞。接着将剩余的红细胞放置于在冰上与溶解液作用,以打破所有红细胞的膜,这样就只留下血红蛋白。而这才是新鲜组织被浸泡了两年的液体。可对于血红蛋白原料如此的制备工作完全不能反映地狱溪岩层里埋藏的实际情况。
6.2
施怀哲等人11用了质谱法(mass spectrometry)鉴定出了恐龙骨细胞的九种肽类(几乎完整的蛋白质),却没有用质谱法去分析这些肽类遇到水解反应的后果,这本很容易做到——因为由于氧化作用,氢氧自由和过氧化物分子对氨基酸(例如天冬酰胺、谷氨酰胺等)的破坏作用会非常明显。另外,他们也该找寻水解反应的证据。既然地狱溪据称曾在内陆浅海的海面之下,因此假如芬顿反应在有水存在的生物软组织中活跃进行,水解作用的破坏力就必然发生。此外,如上所述,芬顿反应牵涉的氢氧自由基和过氧化物对于生物组织的高度破坏作用早已众所周知。10 [14] 因此氨基酸的破坏本应该被观察到。更有甚者,施怀哲等人赤裸裸地宣称,“氧自由基能像甲醛那样促成蛋白质发生交叉连接”,然后她们参考了霍金斯(Hawkins)与戴维斯(Davies)的论文,[15] 可这两人所引用的240篇论文里都证明了氢氧自由基实际上反而对生物组织起破坏作用。纵然有上述提到的一切问题(包括从凝固血液中、血红蛋白问题、血红素分子中释放游离铁的问题、以及芬顿反应产生的氧自由基奇迹般地稳定生物组织的问题),施怀哲等人11却仅用光镜观察了两年后的培养组织,且只发表了光镜照片,而缺乏更具体的研究分析。拉纳1提到了施怀哲仅在部分样本中观察到了以密集浓缩形式存在的铁纤维——而这可联系到为人所众知的生物死亡后在其躯干或四肢低处的血液产物的汇聚现象,这称为尸斑(livor-mortis)。实际上当我在美国显微镜协会( Microscopy Society of America)2013年会议的讲话中,将施怀哲等人论文11里的一张图片上下颠倒反过来放置,并当众发问道,这些铁纤维又怎能穿越大块身体组织并起稳定作用?所有在场者的脑袋都上下点头表示同意。
6.3
将软组织放置于高度人工处理后的血红蛋白里、又在恒温条件下、且放置于实验室容器里达两年、缺乏水分与热量、缺乏蒙大拿州当地融冰与结冰的气象周期变换、也排除了微生物、细菌、植物根系、真菌团、昆虫、啮齿类、加上忽视所有自然环境中其它出现的因素——根本不能代表这些恐龙遗骸埋葬的真实条件。而符合实际情况的实验理当包括所有这些因素。
条件7
拉纳承认高温对软组织的破坏胜于低温,可他却接着声称高温有助于软组织的保存!——拉纳又来这一套了,他到底想说什么东西?蒙大拿的夏天与冬天平均温度分别为31摄氏度(88华氏度)和-13摄氏度(9华氏度),再加上雨水、融雪、结冰等也是不可忽略的环境因素。这就是说从中提取了软组织的三角龙角纵使经历了年复一年的高温与(冰点以下)低温的无数周期变换,依旧展现了惊人的保存状况。
条件8
按拉纳的逻辑,中性酸碱度最适合保存。但高酸度或高碱度不也是如此吗?于是又来这一套了,我又要问拉纳,究竟哪个才是?然而,在地狱溪地层里所有这些生物活动存在的情况下,设想下的中性酸碱度却有助于上文所有这些破坏性生物的生存活动,而这正是三角龙角所处的情况。再一次,这个必要的稳定条件被证明无效。
条件9
最后,拉纳争辩道,处在富含矿物环境里的胶原能保存得更好,是因为这样的环境会包裹胶原并使他们隔离于酶、其它分解者等。然而拥有惊人保存度的骨细胞可不是胶原。我所发现的完全是柔软又灵动的骨细胞,并且数量很多,连矿化的证据都没有。又一次,拉纳勾勒的这个必要条件无效。
结论
无论对于我在蒙大拿洲从离受侵蚀的地面还不到61厘米的开裂三角龙角内剥离出的纤维骨软层,还是从龙角里提取的数千个骨细胞,上述这些条件都不适用。我还可以对他书中的其他部分说更多,但我相信其主张已经被摧毁了。妄图证明铁和芬顿反应可以保存这些惊人细胞的草率实验已经被证明充满了巨大漏洞。而拉纳也不能指望真地存在他所谓的能够保存我们所发现的三角龙角软组织的这些“必要”条件。
鸣谢
我感谢文章的审稿者以及安德鲁·斯奈林(Andrew Snelling)对文稿的改进。
参考与注释
[1] Rana, F., Dinosaur Blood and the Age of the Earth, RTB Press, Covina, CA, 2016.
[2] For example, Mortenson, T. and Ury, T.H. (Eds.), Coming to Grips with Genesis, Green Forest, Master Books, AR, 2008.
[3] Armitage, M.H., Preservation of Triceratops horridus tissue cells from the Hell Creek Formation, MT, Micros Today 24(01):18–23, 2016 | doi:10.1017/S1551929515001133.
[4] 例如, Vardiman, L., Snelling, A.A. and Chaffin, E.F. (Eds.), Radioisotopes and the Age of the Earth: A Young-Earth Creationist Research Initiative, Institute for Creation Research; St. Joseph, MO, and Creation Research Society, El Cajon, CA, 2000; Vardiman, L., Snelling, A.A. and Chaffin, E.F. (Eds.), Radioisotopes and the Age of the Earth: Results of a Young-Earth Creationist Research Initiative, Institute for Creation Research, El Cajon, CA, and Creation Research Society, Chino Valley, AZ, 2005.
[5] Jenkins, J.H., Herminghuysen, K.R. and Blue, T.E. et al., Additional experimental evidence for a solar influence on nuclear decay rates, Astropart. Physics 37:81–88, 2012 | doi:10.1016/j.astropartphys.2012.07.008.
[6] Sibley, A., Variable radioactive decay rates and the changes in solar activity, J. Creation 27(2):3–4, 2013; creation.com/decay-rates.
[7] Armitage, M.H. and Anderson, K.L., Soft sheets of fibrillar bone from a fossil of the supraorbital horn of the dinosaur Triceratops horridus, Acta Histochemica 115(6):603–608, 2013 | doi:10.1016/j.acthis.2013.01.001.
[8] Thomas, B. and Nelson, V., Radiocarbon in dinosaur and other fossils, CRSQ 51(4):299–311, 2015.
[9] Vardiman, L., Snelling, A.A. and Chaffin, 2005, ref. 4.
[10] McCord, J., Iron, free radicals and oxidative injury, J. Nutrition 134(11): 3171S–3172S, 2004.
[11] Schweitzer, M.H., Zheng, W., Cleland, T.P. et al., A role for iron and oxygen in preserving soft tissues, cells and molecules from deep time, Proc. R. Soc. B 281(1775):2013.2741, 2014 | doi:10.1098/rspb.2013.2741; rspb.royalsocietypublishing.org/content/281/1775/20132741.
[12] 详细参见DeMassa, J.M. and Boudreaux, E., Dinosaur peptide preservation and degradation, CRSQ 51(4):268–285, 2015.
[13] Armitage and Anderson, ref. 7, p. 603.
[14] Prousek, J., Fenton chemistry in biology and medicine, Pure Appl. Chem. 79(12):2325–2338, 2007 | doi: 10.1351/pac200779122325.
[15] Hawkins, C.L. and Davies, M.J., Generation and propagations of radical reactions on proteins, Biochim. Biophys. Acta 1504(2-3):196–219, 2001 | https://doi.org/10.1016/S0005-2728(00)00252-8.
