
作者:Yingguang Liu 翻译:流萤
原文发表于Journal of Creation 32(3):7-9.
2018年,斯多克尔(Stoeckle)和泰勒(Thaler)在《人类进化》杂志(Human Evolution)上发表了一篇文章,题为“为何线粒体能定义物种?”(Why Should Mitochondria Define Species?)。1 文章挑战了人们对地球生物历史的认知。两位作者分析了近十年累积的DNA条码资料,得出以下结论:1. 动物界各个物种之间在基因序列上有空白区;2. 现存动物种都是“在最近几十万年内从小群体发展起来的”。该文引起了大众媒体的注意,我们作为创造论者如何看待这些发现?
该基因是否有代表性?
文章使用的DNA序列比较短,本不是用来做谱系分析的,而是用来鉴定物种的,类似于商店里识别商品用的条码,所以叫DNA条码。然而随着DNA条码数据日益增多,斯多克尔和泰勒得以从中寻找谱系发生方面的踪迹。
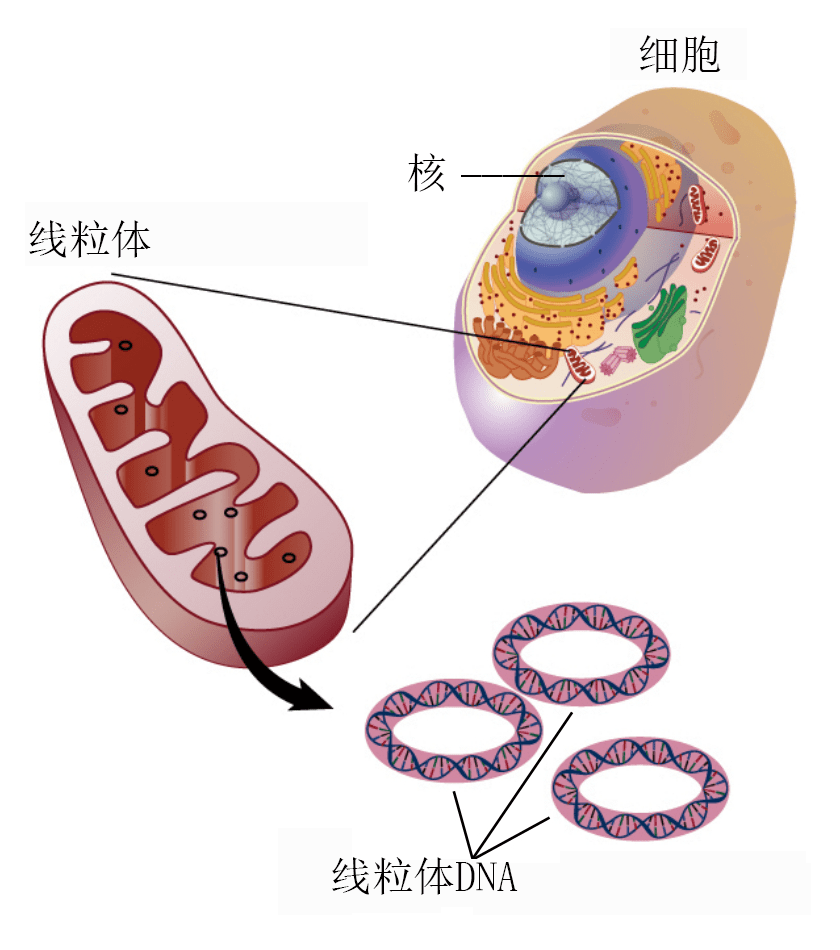
动物DNA条码都取自同一个基因,即线粒体内的细胞色素氧化酶第一亚单位(COI)。细胞色素氧化酶是细胞在燃烧食物的时候直接与氧分子发生作用的蛋白质。虽然动物细胞中 大部分的DNA保存在细胞核内,但是线粒体内也有少量DNA,即线粒体基因组(图1)。该DNA分子是通过母系遗传的,没有父母之间的混合,其序列比核内DNA稳定,而且物种之间也有较强的一致性(同源性),不同物种的COI序列可以用同样的试剂盒来分析。线粒体DNA的同源性也方便了谱系分析。尽管COI基因只占线粒体基因组的5%,不到人体总基因组的百万分之一,但是根据该基因建立的谱系树与根据整个线粒体基因组建立起来的谱系树良好地吻合。根据COI序列进行的动物分类也与分类学专家根据总体性状的分类结果一致。创造论遗传学家纳撒尼尔·詹森(Nathaniel Jeanson)也认为根据线粒体DNA建立的谱系树是准确的、有意义的。2
动物都经历了近期的族群瓶颈?
这篇文章报告的一项重大发现,就是不同物种内部的个体之间在COI基因上的差异都是惊人地相似,平均差异在0.0%到0.5%之间(人与人之间的平均差异是0.1%)。种内变异主要是突变造成的。斯多克尔和泰勒认为这些突变并不影响繁殖能力(中性突变),不受选择,所以能够随时间而累积。线粒体基因的突变比核内基因突变更不容易被自然选择所识别,因为线粒体内的基因读码系统比较简单,同义tRNA比较少,而且没有选择性剪接。无论这些突变是中性还是近中性,自然选择都不会终止突变的累积。3
随机突变的累积可用作一种分子钟。虽然历史上不同物种的突变率可能有所变化,但是突变的数目还是和时间相关的。既然所有的动物种群都累积了相同数目的突变,这就强烈提示所有动物群体都是在同一时间内发展起来的。所以,斯多克尔和泰勒得出如下结论:
“就是说,现存的动物族群,不管它们目前的规模有多大,也不管它们与哪一个时代的化石相似,都是在过去20万年以内发展起来的。20万年前,线粒体DNA没有种内差异。”
这两位作者提议,线粒体DNA的种内一致性可以用“瓶颈、始祖效应、世系重整或基因取代”来解释。所谓“世系重整”,就是一种基因形式通过随机漂移排除了其他形式(类似于人类社会中某些姓氏因偶然因素而消失),“基因取代”则是强力的正向选择的结果。“世系重整在小群体中的效果最强”,而基因取代则需要“群体中所有的线粒体基因组都从同一个母亲重新发源。”
然后作者们推测:
“不知道是否发生了一系列的事件,为动物界各个物种带来了类似的后果,还是只发生了一个事件,影响了整个动物界。根据简约原则(奥卡姆剃刀),应该只用一个过程来解释。”
同样是这两位作者,在几年以前还反对这种想法:
“如果是瓶颈限制了变异,而所有动物的历史都不长,那就意味着不久前所有的物种都经历了群体数量的锐减。这不太可能——几乎就是诺亚方舟假说——尽管长时期气候变化循环也可能造成广泛的周期性瓶颈。”4
他们已经捅开了这层窗纸!数据完全符合年轻地球和全球性洪水,尽管还不能区分这是创造的始祖效应和洪水的瓶颈效应,因为创造和洪水之间的时间并不长。
时钟可靠吗?
至于线粒体时钟在历史上是否恒速运作,线粒体DNA的突变率具体是多少,我们无从知晓,但是我们可以根据作者估算时间的方法对这篇文章所给出的时间跨度做出评价。既然现存人类和动物群体是在同一时间发源的,斯多克尔和泰勒便根据公认的人类历史来估算动物的历史,而人类历史又是基于“数千人的基因组分析、数万个线粒体基因序列、古生物学、人类学、历史学和语言学”。化石(古生物学)在时钟校对中起了重大作用,5,6 但是对化石的年代争议很多。相反,詹森只考虑目前观察到的线粒体DNA突变率(每代每人5-8个突变),发现人类线粒体基因组的变异情况更符合圣经记载的六千年左右的历史,而不是公认的20万年。7,8
林奈和达尔文
斯多克尔和泰勒的另一个重要发现是动物族群之间缺乏基因序列上的连续性。虽然种内个体之间的变异很小,但是种间的序列差异却很大。他们指出:
“DNA条码的分布有两个重要特点:1. 群内变异低;2. 群间存在序列上的空白区,没有过渡序列。”
这本不足为奇,因为条形码就是用来区别物种的。如果物种之间在序列上有严重重叠,COI基因就不能当条码用了。然而斯多克尔和泰勒强调这种不连续性是很有道理的。分子谱系分析本来就是要把物种间的相似性和差异性做定量分析,而DNA和蛋白质序列一直给人一种印象,就是种群之间只有量的差异。与宏观性状不同,人们很难在分子序列中看出质的区别。由于今天大多数谱系树都是基于序列比较,分子进化论对其有强烈的依赖性,尽管这些谱系树与古生物学分类会有冲突,不同的分子谱系树之间也会相互矛盾。9 然而随着DNA条码资料的大量积累,斯多克尔和泰勒却看出了线粒体基因组的种内变异被“限制在狭窄的范围内”。
他们正确地返回到林奈与达尔文之争,这两位作者引用艾夫斯(Aves)等人的说法:
“艾夫斯和他的同事们在关于谱系地理学的一篇开创性著作中,提到了生物学上的一个长期争执,即林奈学派与达尔文学派之间的争执,林奈学派认为物种是各自独立的,而达尔文学派则强调种内的渐进性变化产生了新物种。”10
卡尔·林奈(Carl Linnaeus, 1707-1778)从亚里士多德时代留传下来的“自然等级”出发,撰写了他的《自然系统》,开创了生物分类学和双名法,强调物种分别,物种间有不可逾越的鸿沟。同时代的布封(Comte de Buffon, 1707–1788)则反对这一观点,撰写了《自然史》,倡导自然界有“看不见的过渡”。11 关于物种间是连续还是断续的争论一直延续到今天。斯多克尔和泰勒指出:
“种内条码紧凑成簇,种间却一片空白,这些都是动物界的关键事实,进化论必须予以解释。”
他们甚至为此提出了一个假说:
“最亲近的现存物种之间的距离或大或小,这可能反映了不同数量的已经灭绝的过渡序列。”(下划线是本作者加上的)
进化论的新定律?
斯多克尔和泰勒认为空白区可以用灭绝来解释,他们引用了1973年范瓦伦(Leigh Van Valen)提出的“进化论的新定律”,又叫“灭绝定律”。范瓦伦写道: “现有资料表明,所有的族群都以特有的、恒定的速度走向灭绝。” 12
这一定律主要是基于化石记录。按照范瓦伦的说法,灭绝的主要原因是种间的生态互动,所以灭绝的概率与物种的年龄无关。范瓦伦和他同时代的多数人一样,相信新物种在不断地出现。
既然没有一个物种能长期生存,那么从历史上的任何时侯去看,所有的物种必然都是新的。然而,该定律认为不同种群走向灭绝的速度不同,所以不能解释为什么现存动物种群的线粒体年龄都差不多。
值得指出的是,灭绝定律得到了当代生物学的支持。植物遗传学家圣弗德(John Sanford)在他的《基因熵》一书中,提出所有的物种都有一定的寿命,不是由于外在因素,而是由于基因组因随机突变的累积而最终瓦解。3 圣弗德不认为新的生命形式会自然发生,所以整个生物界的历史也不是很长。
下一步
斯多克尔和泰勒指出,还有23个小型无脊椎动物门,其DNA条码的分布尚不明了。随着条码分析资料的日积月累,它们的历史轨迹也会越来越清晰。
创造论生物学家一直试图为圣经上讲的“受造类”做出明确的定义。这些“类”是现存的和已经灭绝的一切物种的祖先。斯多克尔和泰勒的研究虽然显示多数动物属内缺乏种间连续性,但是也有一些个别的属,比如熊属,种间变异相对不大。这不难解释,熊属内的各个物种可能都是同一个受造类的后代,至今种间还可以杂交。或许DNA条带数据可以帮助我们鉴定受造类并了解大洪水前后受造类的历史。
结论
斯多克尔和泰勒之所以能一投石激起千层浪,是因为他们所发现的事实挑战了统治学术界多年的达尔文渐进主义思想。虽然动物种群的绝对年龄值得讨论,但是普遍年轻的线粒体基因是与近期创造或者大洪水所造成的基因瓶颈一致的。这些发现不仅不支持漫长进化的信条,而且DNA序列数据的不连续性也驳斥了新达尔文主义者一贯倡导的突变过渡机制。
参考文献
- Stoeckle, S.M. and Thaler, D.S., Why should mitochondria define species? Human Evolution 33(1–2):1–30, 2018. 10.14673/HE2018121037.
- Jeanson, N., Replacing Darwin: The new origin of species, Master Books, Green Forest, AR, pp. 248–249, 2017.
- Sanford, J.C., Genetic Entropy and the Mystery of the Genome, FMS Publications, Waterloo, NY, pp. 45-68, 2008.
- Stoeckle, M.Y. and Thaler, D.S., DNA barcoding works in practice but not in (neutral) theory, PLOS ONE 9:7e100755, 2014.
- Cann, R.L., Stoneking, M., and Wilson, A.C., Mitochondrial DNA and human evolution, Nature 325(6099):31–36, 1987.
- Stringer, C., The origin and evolution of Homo sapiens, Philosophical Transactions of the Royal Society B: Biological Sciences 371(1698):20150237, 2016.
- Jeanson, ref. 2, pp. 182–186, 2017.
- For a recent summary of molecular genetic evidence towards a young creation see: Tomkins, J., Empirical genetic clocks give biblical timelines, J. Creation 29(2):3–5, 2015.
- Tomkins, J. and Bergman, J., Evolutionary molecular genetic clocks—a perpetual exercise in futility and failure, J. Creation 29:26–35, 2015.
- Avise, J.C. Neigel, J.E., and Arnold, J., Demographic influences on mitochondrial DNA lineage survivorship in animal populations, J. Molecular Evolution 20:99–105, 1984.
- Sloan, P.R., The Buffon–Linnaeus controversy, Isis 67(3):356–375, 1976.
- Van Valen, L., A new evolutionary law, Evolutionary Theory 1(1):1–30, 1973.
