
作者 唐·巴顿 发表日期:2013年11月26日
最后修订于2018年5月17日
引言
生命的起源也称为生命的自然发生,有时也称作化学进化。生命的基础是富含信息的长链分子,例如DNA和RNA。这些分子中带有制造蛋白质的指令,而蛋白质则是生命的砖瓦。但要读取DNA / RNA中的信息以制造蛋白质,要复制DNA或RNA以制造新细胞(繁殖是生命的标志),都需要大量的由DNA / RNA编码的蛋白质。因此,在生命一开始,就必须同时有DNA / RNA和蛋白质:这是生命自然发生假说一个无法解决的“先有鸡还是先有蛋”的难题。
因此,对于那些坚持仅仅通过自然的过程(物理和化学)就能产生生命的人来说,生命的起源是一个棘手的问题。
一些进化论者声称生命的起源不是进化的一部分。 然而,几乎每一本进化生物学教科书都有关于生命起源的章节。加州大学伯克利分校的“进化 101”课程中包含生命起源的内容,其标题为“从原始汤到细胞——生命的起源”。 1 “万物源于进化”理论的高调捍卫者们,例如迈尔斯(P.Z. Myers)和尼克·马茨克(Nick Matzke)等人都同意生命的起源是进化的一部分,理查德·道金斯(Richard Dawkins)也是如此。2
之前著名的进化论者却吉 (G.A. Kerkut)确曾将进化论分为两种:包含生命起源的广义进化论(General Theory of Evolution,GTE),以及仅涉及生命多样化的狭义进化论(Special Theory of Evolution,STE),后者据称是达尔文1859年的著作,《物种起源》,的主题。3
直到最近,一些进化论的捍卫者才试图将生命起源与进化分开。 这可能是因为近来的生命科学研究,即使在最简单的活细胞中,也发现了一个又一个复杂的分子机器,使得自然主义(生命自然产生)的问题变得越来越困难,解决这些难题的希望正在迅速消失。
那么,生命的产生需要什么? 为了通俗地解释生命起源相关的内容,我们在这里将生命起源的问题分解为许多子话题(尽管这仍然可能是比较烧脑的)。
 形成活细胞需要什么材料? 活细胞能够从周围环境中获取所需的所有资源并自我复制。 第一个细胞必须能够独立生存; 也就是说,它不能依赖于其他细胞而存活,因为其他细胞不存在。 寄生生物不能用作“第一个生命”的模型,因为它们需要其他的细胞才能存活。 基于这个原因,病毒之类也不可能是“生命的先驱”,因为它们必须寄生于活细胞内才能进行自我复制。朊病毒,致病性畸形蛋白质,同样与生命起源无关,因为它们的“复制”不过就是让宿主细胞产生的蛋白质变形。
形成活细胞需要什么材料? 活细胞能够从周围环境中获取所需的所有资源并自我复制。 第一个细胞必须能够独立生存; 也就是说,它不能依赖于其他细胞而存活,因为其他细胞不存在。 寄生生物不能用作“第一个生命”的模型,因为它们需要其他的细胞才能存活。 基于这个原因,病毒之类也不可能是“生命的先驱”,因为它们必须寄生于活细胞内才能进行自我复制。朊病毒,致病性畸形蛋白质,同样与生命起源无关,因为它们的“复制”不过就是让宿主细胞产生的蛋白质变形。
首先需要的是合适的材料。这有点像做蛋糕; 如果没有香蕉或面粉,你就不能做出香蕉蛋糕。
备齐合适的材料
“生命起源于化学汤”的假说中存在一个主要的问题:所有的生命材料必须存在于同一个地方,这样才有可能组装出一个活细胞。 但生命所必需的化学材料具有羰基(> C = O),羰基化合物与氨基酸和其他氨基(-NH 2)化合物会发生破坏性的反应。含羰基的分子包括“糖”,4 糖也是构成DNA和RNA的骨架。 在活细胞中,羰基化合物与氨基化合物被分隔在不同的区域内保护着,以防止这种交叉反应,有些细胞可以在交叉反应发生后进行修复,但是化学汤没有这种保护或修复机制。
细胞是由较简单的化学物质组成的极其复杂的结构。本文不打算详述第一个细胞所需要的每种化学物质; 这需要一本书的内容或更多的篇幅才能说清。本文重点介绍在任何生命起源模式中都必须存在的一些最基本的生命材料。
a. 氨基酸
生物体中充满了蛋白质,蛋白质是由线性串联的氨基酸组成的。例如,酶是特殊的蛋白质,用于催化化学反应(催化剂)。 比如在我们的唾液中的淀粉酶,可以将大米、面包、马铃薯等食物中的淀粉分子分解成较小的分子,然后这些小分子可以被进一步分解成葡萄糖分子(淀粉分子的组成单元)。我们不能吸收淀粉,但我们能够吸收葡萄糖并将其作为能源来驱动我们的身体。
生命所必需的一些反应在没有酶的条件下进行得非常缓慢,即使有数十亿年的时间,也不能产生足够的产物以供生命的需要。5
其他蛋白质组成了肌肉、骨骼、皮肤、毛发以及细胞和身体的各种结构部分。人类可以生产超过100,000种蛋白质(还可能是数百万种,没有人确切知道是多少种),而典型的细菌可以产生超过一两千种不同的蛋白质。
 蛋白质由20种不同的氨基酸组成(某些微生物拥有一种或两种额外的氨基酸)。 氨基酸不是简单的化学物质,如果没有酶(酶本身也是由氨基酸组成的),它们就不容易正确地建造,见图1。
蛋白质由20种不同的氨基酸组成(某些微生物拥有一种或两种额外的氨基酸)。 氨基酸不是简单的化学物质,如果没有酶(酶本身也是由氨基酸组成的),它们就不容易正确地建造,见图1。
几乎每一本生物学教科书仍然在介绍1953年发表的米勒-尤里(Miller-Urey)实验,这个实验在没有酶的条件下制造了一些氨基酸,因此它通常被解释为“生命起源”的模拟实验,但这种解释要么是非常无知,要么是存心欺骗。
虽然这个实验制造了微量的正确氨基酸,但是这个实验所设定的条件从来没有在地球上发生过。例如,烧瓶中的“大气”中如果含有微量氧气,就不能合成任何物质。另外这个实验还同时制造了一些错误类型的氨基酸,以及其他会发生“交叉反应”的化学物质,它们会妨碍任何有用物质的形成。
在自然界中,功能性蛋白质所需的氨基酸绝不可能通过这个实验所模拟的条件来制造。6 当斯坦利·米勒(Stanley Miller)在1983年用一种稍微更现实的气体混合物重复做这个实验时,他只得到了微量的甘氨酸,这是制造蛋白质所需的20种氨基酸中最简单的一种。7
如何才能得到正确氨基酸的混合物仍然是一个未解之谜(而且还有另外一个主要的问题,请看下面“手性”一节)。
b. 糖
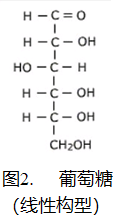 有些糖可以仅通过化学方法制造,而不需要酶的参与(请记住,酶只能由细胞制造)。然而,在没有酶参与的条件下制造糖需要一个碱性的环境,而在这种环境下氨基酸是不能合成的。
有些糖可以仅通过化学方法制造,而不需要酶的参与(请记住,酶只能由细胞制造)。然而,在没有酶参与的条件下制造糖需要一个碱性的环境,而在这种环境下氨基酸是不能合成的。
在合成糖的化学反应过程中,要求环境中没有含氮化合物,如氨基酸,因为它们会与制造糖的中间产物——甲醛,以及糖发生反应,产生出没有用(非生物)的化学物质。
核糖构成RNA的骨架,核糖经改造后构成DNA的骨架,二者都是所有活细胞的重要组成部分。核糖在化学进化中是一个难题。因为在现实世界中,在接近中性pH(既不酸性也不碱性)的环境中,核糖是一种不稳定的糖(它会快速地分解),半衰期太短。8
c. DNA和RNA的部件
在没有来自活细胞的酶的帮助下,我们怎样才能得到DNA和RNA中的化学“字母”——核苷酸呢?在没有酶参与的条件下,使用化学方法制造核苷酸,就需要甲醛(H 2 C = O)与氰化氢(HC≡N)进行反应。 然而,甲醛和氰化物都是致命的剧毒化学品(特别是后者)。它们将非常严重地破坏重要的蛋白质(如果有的话)!
 胞嘧啶(图3)是DNA和RNA中5种必需的核苷酸碱基中的1种,在生命产生之前的任何现实的情景中都很难制造,也非常不稳定。8
胞嘧啶(图3)是DNA和RNA中5种必需的核苷酸碱基中的1种,在生命产生之前的任何现实的情景中都很难制造,也非常不稳定。8
DNA和RNA都具有糖和磷酸基团交替连接构成的主链骨架。上面已经讨论了制造糖的问题。磷酸离子会被海水中丰富的钙离子所沉淀,或粘附在粘土颗粒的表面上,这两种情况都会阻止磷酸被用来制造DNA。
d. 脂类
脂类(“油脂”)对于形成细胞膜是必不可少的,后者包裹细胞内容物并执行其他细胞功能。细胞膜由几种不同的复杂脂类组成,是独立生存、自我繁殖的细胞的重要组成部分。
脂类比糖或氨基酸具有更高密度的能量,因此它们在化学汤中的形成是任何生命起源理论的一个难题(从热力学的角度来看,含有高能量的化合物比低能量的化合物更加难以形成)。
即使假设没有氧气存在(在一个“还原性”的大气中),作为所有细胞膜的主要成分的脂肪酸也是很难生产的。即使产生了脂肪酸分子,生命所必需的离子,如镁和钙离子(每个原子具有两个电荷,2+,即二价),也会与脂肪酸结合,并使脂肪酸沉淀而失去作用。9 这如同在(富含镁和钙离子的)硬水中使用肥皂(主要成分是脂肪酸盐)进行清洗,就会发生相同的沉淀反应,形成“浮渣”。
有些自然发生假说的倡导人喜欢画一个空心脂球(囊泡)的示意图,这样的简单结构在一定的条件下可以在试管中形成。然而,这种“膜”永远不会形成活细胞,因为细胞需要通过脂膜向内和向外运送物质。这种进出细胞的运输需要非常复杂的“蛋白质—脂质”复合物,称为转运通道,其运行方式类似于电动机械泵。出入细胞的各种化学物质都有自己的特异性通道(好比用于抽水的泵不一定适合用于抽油)。许多泵能够借助于高能化合物(如ATP)沿着与自然梯度相反的方向(从低浓度到高浓度)进行主动运输。即使在顺着自然梯度(从高浓度到低浓度)进行运输时,也通常需要载体蛋白来促进。
细胞膜还维持着细胞内酶活性所需的稳定的pH,并保持着各种矿物质的恰当浓度(例如钠不能太多)。 这需要运输通道(“泵”)在细胞的控制下特异性地运输氢离子(质子),这些泵具有很高的特异性。 10
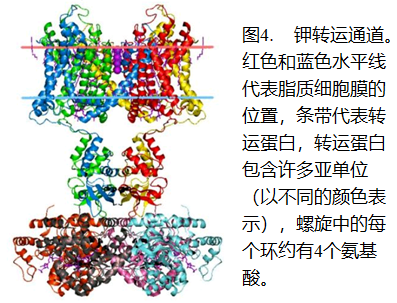 细胞膜的运输是如此重要,以至“大多数生物的基因组中,20-30%的基因所编码的都是细胞膜蛋白”。11已知最小的自主生存的生物,寄生性生殖支原体的基因组,在其482个蛋白编码基因中,26个基因编码细胞膜运输蛋白 12 。
细胞膜的运输是如此重要,以至“大多数生物的基因组中,20-30%的基因所编码的都是细胞膜蛋白”。11已知最小的自主生存的生物,寄生性生殖支原体的基因组,在其482个蛋白编码基因中,26个基因编码细胞膜运输蛋白 12 。
纯脂质的膜甚至不允许带正电荷的金属离子(例如钙、钾、镁、铁、锰等)或带负电的离子(如磷酸基、硫酸基)被动运动出入细胞,而这些都是生命必需的营养物质。纯脂质的膜会排斥这些溶于水而不溶于油脂的带电离子。事实上,一个简单的脂肪膜还会阻止水的通过(试一下将橄榄油和水混合在一起)!
膜转运蛋白是活细胞所必需。
在20世纪20年代,生命起源于肥皂泡(脂肪球)的理论很受欢迎(奥帕林的“凝聚”假设),但那时候人们还未知道生命中DNA和蛋白质合成的极度复杂性,也不知道细胞膜奇妙的功能。这些假说极端天真,但是相关的视频今天仍然可以在YouTube见到,显示脂质泡,甚至还能分裂,好像这就能解释生命的起源(见视频:能自我创造的细胞? 当然不是! )。
e. 手性
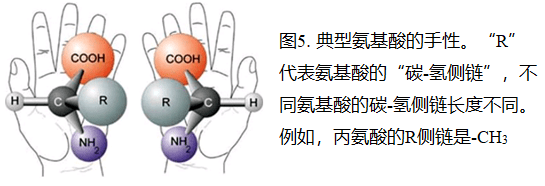 氨基酸、糖和许多其他生物化学物质都有三维结构,通常有两种彼此互为镜像的结构;像你的右手和左手是彼此互为镜像。这被称为手性(图5)。
氨基酸、糖和许多其他生物化学物质都有三维结构,通常有两种彼此互为镜像的结构;像你的右手和左手是彼此互为镜像。这被称为手性(图5)。
现有的生物体内的生物化学物质都是由一种手性结构成分构成的(纯手性):例如所有的氨基酸都是左手性(左旋),所有的糖都是右手性(右旋)。这就产生了一个问题,在没有酶参与的化学反应中(如米勒-尤里实验),当其生成氨基酸时,会同时产生右手性和左手性氨基酸的混合物。化学方法(例如甲酸反应)合成的糖也一样。13
研究生命起源的科研人员一直在设法对付这个问题。他们提出了各种可能的方案,但此问题仍未得到解决。14即使通过一些完全人工的方法,获得了99%手性纯度的氨基酸(这通过“自然”的机制不可能获得),也不能解决这个问题。生命需要100%纯左手性的氨基酸。原因是:若是蛋白质中的一个左手性氨基酸被右手性氨基酸替代了,就会使蛋白质产生不同的三维形状。而要得到生命所必需的蛋白质,这种情况一次也不允许发生。
细胞存活的最低要求是什么?
独立生存的细胞若要使用从周围环境中获得的化学物质和能量以制造细胞部件并能自我复制,就必须具有:
- 细胞膜:该结构能将细胞与环境隔开。细胞膜必须能够维持细胞内与细胞外不同的化学环境(如上文所述)。若没有细胞膜,生命的化学过程是不可能进行的。
- 一种存储信息或遗传指令的方法,来指导细胞如何复制出另一个细胞(繁殖)以及在不同的情况下如何进行细胞内的运作。唯一已知的信息保存方法是使用DNA,任何使用其他材料(例如 RNA)的提议尚未被证明是可行的——如果使用其他材料,那就还需要设法将该系统中的信息转化为DNA,后者是所有已知生命的基础。15
- 读取(2)中的信息,并在合适的时间、按照适当的数量制造细胞的构成材料。细胞的主要的构成材料是蛋白质,它是由数百至数千个大概20种不同的氨基酸分子聚合串联成的大分子(聚合物)。从DNA指令中制造蛋白质的唯一已知的(或甚至可想到的)方法,需要100多种蛋白质和其他的辅助因子共同参与,至少包括:
- 纳米机器,例如RNA聚合酶(已知最小的RNA聚合酶大概含有4,500个氨基酸)
- 旋转酶:增加或解除DNA的螺旋,使DNA中的信息能够被“读取”(这是些体积很大的蛋白质)
- 核糖体:制造蛋白质的亚细胞“工厂”
- 至少20种转运RNA分子,这些转运RNA分子可以选择正确的氨基酸,将其按照DNA上指定的顺序排列。已知所有的细胞至少有61种转运RNA分子,因为大多数氨基酸对应着不止一种DNA密码(每个密码由DNA上的三个字母组成)。 转运RNA具有根据DNA密码来确保选择正确氨基酸的复杂机制。
- 还需要一些机制以确保所制造的蛋白质以正确的方式折叠成正确的三维结构,包括以分子伴侣来保护蛋白质避免错误的折叠,还有称为伴侣蛋白的折叠“机器”,新制造的蛋白质会进入伴侣蛋白中,使新造的蛋白能够进行正确的折叠。所有细胞都有这些机制。
哇!上述只是最基本的机制。
(这是一个简要的蛋白质合成动画,其中包括RNA聚合酶、核糖体、转运RNA、伴侣蛋白和分子伴侣的作用。所有活细胞都具有这种蛋白合成系统。来源: youtube)
- 需要一种从环境中的简单化学物质制造细胞所需要的生化分子的方法。这包括制造ATP,ATP是生命的通用能量货币。已知所有的活细胞都有ATP合成酶,一种非常复杂和高效的电动旋转马达,可以制造ATP(这种马达也可以反过来使用ATP产生电流,以驱动细胞内/外的其他反应和运动)。
- 需要一种复制信息并将其传递给后代的方法(繁殖)。最近有人模拟了已知最简单的能独立生存的细菌的一次细胞分裂过程(这个细菌“只有”525个基因),模拟实验需要128台台式电脑一起工作10个小时。16
以上给出一些提示,以理解第一个活细胞的生存需要什么前提。
几年前有人启动了一个有趣的研究项目,要确定能独立存活的最简单细胞(就是一个不依赖于另一个生物体就可以生存的最小的细胞)是什么样的。然而,这个细胞还是需要富含营养的培养基来为它提供大量复杂的有机化合物,因此它所需要的许多生物化学物质都不必自己合成。现在已知这个最简单的细胞也需要超过400种蛋白质和RNA零件,17当然,这意味着它的DNA需要含有制造这些蛋白质和RNA的指令。也就是说,它的DNA需要含有超过400个“基因”。我们稍后会继续讨论这个问题。
聚合物的形成(聚合反应)
生命不仅仅由氨基酸或糖组成,而且还需要很多聚合物,它们是由较简单的化合物以串或链的方式连接在一起的。多糖是糖的聚合物;蛋白质是氨基酸的聚合物;DNA和RNA是核苷酸的聚合物。多糖是三者之中最简单的,其链中连接着的通常都是相同的糖分子,如葡萄糖在植物中聚合成淀粉,在动物中聚合成糖原。蛋白质比多糖复杂得多,其链中的每个氨基酸可以是20种不同的氨基酸中的任何一种。DNA和RNA链由四种不同核苷酸连接而成。
另一方面,水是活细胞的必需成分,典型的细菌中的水占其总质量约75%。水作为“通用溶剂”,是细胞各种成分的必要载体,是一切生命活动的环境。
这就为生命起源理论带来一个极大的难题。比如,当2个氨基酸分子聚合时,会释放一个水分子。这意味着在有水的情况下,水会推动逆向反应,造成聚合物的分解(水解);也就是说,除非水能被主动去除,否则蛋白质会分解而不会形成。细胞之所以能克服这一难题,是靠着保护核糖体内的反应位点,借助于能量不让水进入反应位点,并提供能量来驱动聚合物的形成。 因此,对于所有的生命起源理论而言,如何才能形成长度超过几个氨基酸的蛋白质是极为困难的问题(再多的时间也不能解决这个问题,延长时间只能导致更多蛋白质被水解)。
聚合物的形成还需要被连接在一起的每一个零件(单体)都具备两个官能团。这就意味着用于制造蛋白质的氨基酸(或用于制造多糖的单糖)至少具有两个活性位点,以允许另一个氨基酸(或单糖)连接到它的两侧。形成蛋白质的氨基酸须具有至少一个氨基(-NH 2)和一个羧基(-COOH),每一个氨基酸的氨基与另一个氨基酸的羧基连接,从而使氨基酸链不断延长。仅具有一个活性位点(单官能团) 的化合物会终止聚合物链的形成。生命起源理论的问题在于,任何可能产生氨基酸的化学反应,也会产生单官能团的物质,后者会阻止蛋白质的形成。18
核酸(如DNA和RNA)的主链骨架是糖的聚合物。同样,如果存在一些单官能糖就会终止DNA / RNA的形成,而且在有水的环境中,水分子会促进DNA / RNA的分解,而不是聚合。
生命的起源是编程问题,而不仅仅是化学问题
 上述资料足以完全否定自然主义的生命起源理论,但我们还未提及最重要的问题,即程序的起源。生命不仅仅依赖于聚合物,而且更依赖于聚合物中亚单位的特定排列方式; 氨基酸的特定排列产生具有功能的蛋白质/酶;核苷酸碱基的特定排列产生具有功能的DNA和RNA。
上述资料足以完全否定自然主义的生命起源理论,但我们还未提及最重要的问题,即程序的起源。生命不仅仅依赖于聚合物,而且更依赖于聚合物中亚单位的特定排列方式; 氨基酸的特定排列产生具有功能的蛋白质/酶;核苷酸碱基的特定排列产生具有功能的DNA和RNA。
亚利桑那州立大学“超越科学基本概念中心”主任、天体生物学家保罗·戴维斯(Paul Davies)说:
“要解释生命是如何开始的,我们需要了解生命独特的信息管理是如何产生的。“
“生命管理信息的方式有一种逻辑结构,这与单纯的复杂化学有着根本的区别。因此,单纯的化学过程并不能解释生命的起源,就好比对硅、铜和塑料的研究并不能解释电脑是如何运行程序的。”19
戴维斯明确清晰地提出了这个问题,应该不会让他的进化论同事感到惊讶,因为这与他十多年前的公开言论很相似。例如“真正神秘的是活细胞中的软件,而不是其硬件。” 他还说:”愚蠢的原子何以自发地编写自己的软件呢? …… 没人知道… …”。20
任何试图解释生命自然发生的理论,若没有解释信息处理系统是如何起源的,若没有解释记录在活细胞DNA里的信息,那都是在回避问题。我们只需要看看最简单的能独立生存的细胞就可以断定,在仅依赖于物理和化学过程的理论里(不允许智能设计),信息的起源是不可能解决的。
二十世纪最杰出的科学哲学家之一,卡尔·波普尔(Karl Popper)爵士意识到:
“生命和遗传密码是怎么样自然产生的是一个令人不安的谜团,其原因是:遗传信息没有任何生物学功能,除非它被翻译;也就是说,除非它其中的遗传信息指导了蛋白质的合成,即蛋白质的结构由遗传信息确定。 但是……细胞(至少是现代细胞——也是我们所唯一知道的)翻译遗传信息所需要的分子机器至少由50个大分子成分组成,而这些大分子本身也编码在DNA中[编辑补充:我们现在知道,至少需要由100个大分子组成这些机器]。因此,若不是使用某些翻译产物,就不能翻译遗传密码。这构成了一个令人困惑的循环; 对于任何关于遗传密码起源的模型或理论,这都似乎是一个真正的死循环。”
“因此,我们或许要面对一个可能性,即生命的起源(就如物理学的起源一样)成为科学上一个不可逾越的界限;是在一切试图将生物学还原为化学和物理学的努力之后,最终残余的一个问题。” 21
DNA密码的起源
波普尔所描述的DNA信息存储系统不能源于化学过程,而必须有一个智慧的来源。22 如果我们考虑一下其他的编码系统,例如摩尔斯电码或书面的字母信息,其中的符号用于表示语言的发音,这种编码系统只能来源于智慧。例如,“a”在英语中通常如它在“cat”中的发音,这是一个随意形成的惯例;字母的形状并没有表明它应该如何发音。同样,从物理和化学定律解释DNA编码系统是不可能的,因为代码和代码记录的信息之间没有物理或化学关系。
用DNA序列来编码蛋白质可以有亿万种方式。任何一套DNA密码的起源都是一个大难题,更何况现有的遗传密码恰恰是“达到或非常接近于错误最小化的最优总体方案,即所有可能的代码方案中最好的。”23 这是因为,用64个“密码子”24来编码20种氨基酸,几乎所有的氨基酸都对应不止一个密码子(有几个常见的氨基酸,如亮氨酸,有六个密码子)。25 这些多重密码子有时被称为“重叠”,通常被认为是“超出了需要”或者“多余”。 然而,额外的密码子是经过优化的,使得DNA编码中最容易发生的单字母错误(突变)一般地不至于造成氨基酸水平上的改变;即使有改变,也是变成化学结构相似的氨基酸(因此对制造出的蛋白质的结构破坏性不大)。
通过额外的密码子还能进行“翻译水平控制”,以实现蛋白质合成数量的复杂控制。该控制系统在细菌和高等生物中都会起作用。26
密码系统不可能分阶段出现,逐步优化。假设通过某种令人难以置信的侥幸,出现了一个可以运行的编码系统,那么在此系统出现之后,基本代码就不会发生重大的变化,因为代码和解码系统(读码机器)必须同时改变,(例如,在一些细菌的基本代码中存在一个非常微小的变化,即使用三个正常的“终止”密码子中的一个,编码了一个额外的氨基酸20)。 所以除非在生命出现之初又发生了另一个难以置信的“自然”侥幸事件,最优化的密码系统无法解释。
除了编码系统,还有信息
我们不仅需要解释信息存储系统是怎么来的,还要解释存储在DNA上的蛋白质信息或指令。我们再来看最简单的细胞,科学家通过从自主生存的微生物中敲除基因来确定哪些是“必需的”,得知这种最简单的细胞需要超过400种蛋白质和RNA零件。所有这些的信息必须在DNA上编码,否则假想中的第一个细胞就不会制造这些零件,更不能自我繁殖以产生另一个细胞。将DNA的四个“字母”所编码的、最简单的细胞所必须的信息打印出来,会是一本很厚的书。
按照保罗·戴维斯的比喻,遗传信息问题类似于电脑的程序。我们如何解释电脑程序的存在? 首先要有编程语言(Python、Fortran、C ++、Basic、Java等),然后是用该编程语言撰写的一套实际指令。DNA起源的问题同样是双重的:编程语言的起源和程序的起源。
从更简单的东西“进化”到最简单的细胞,如此提议的人需要显示从他们假设的更简单的东西进化到第一个活细胞的途径。热衷于生命自然发生假说的人经常求助于“数十亿年”作为解决问题的忽悠方法,但并不给出进化的机制。即使加添再多的时间,朝着错误方向进行的反应也不会逆转,不会自己朝着正确的方向掉头。
生命也需要纠错系统
分子生物学揭示,即使是最简单的细胞也是极度复杂和精妙的。如上所述,信息存储在DNA中,然而DNA是一种非常不稳定的分子。 一份报告说:
“‘人们普遍认为DNA是坚如磐石的——非常稳定’,范德堡大学(Vanderbilt University)的生物科学副教授勃兰·艾希曼(Brandt Eichman)指出,‘但实际上DNA反应性很高。在普通的一天,人体细胞内的DNA中约有一百万个碱基被破坏。’”27
因此,所有细胞必须具有纠正DNA结构改变或编码信息错误的系统。如果没有这些纠错系统,DNA序列中的错误就会累积并最终导致细胞死亡(称为“错误灾变”)。所有活细胞都有这一特性,这为生命起源理论雪上加霜。
假设原始汤中出现了一个带有信息的DNA分子,这个DNA分子必须被准确地复制,否则信息会因复制错误和化学损害而丢失。如果没有有效的DNA修复机制,遗传信息将迅速被降解。 然而,构建这种修复机器的指令是在它要修复的DNA分子上编码的,这是生命起源理论的另一个死循环。28
当科学家发现生活在极端条件下的细菌时,例如海洋中的深海热液喷口,喷口附近的细菌被称为“原始生命”,因为一些研究生命起源的人提出生命可能是在这些地方开始的。 然而,这些被称为“极端微生物”(因为喜欢极端环境)具有相当复杂的DNA纠错系统。例如,耐辐射奇球菌是一种可以耐受极端剂量电离辐射的细菌,这个剂量会杀死你、我或其他细菌。奇球菌的DNA也会受到损伤,甚至会被打成许多碎片。但是在损伤后的数小时内,细菌能激活大约60个基因来修补断端,重建基因组。29
热液喷口是不适宜生存的高温环境,生活在那里的微生物的DNA不断受到破坏,因此它们必须具有复杂的DNA保护和纠错系统才能生存。它们一点都不简单,对于生命起源也没有提供任何可行的模型。30
此外,所有细菌,不仅仅是“极端微生物”,都必须拥有复杂的纠错系统,这些系统涉及许多基因,当纠错系统因突变而失效时,细菌就变得无法生存。 这又为生命起源提出了另一个难题。
生命起源的过程
生命是起源于一个温暖的池塘(如达尔文所推测的)、深海热液喷口附近、粘土颗粒表面、还是在别的地方以其他方式起源的?这么多种环境被提出来,但又没有定论,就表示这些环境都存在重大缺陷。
温暖的池塘和深海热液喷口的一个主要问题是水的存在,水会阻碍许多必需的反应,如聚合反应。此外,深海热液喷口的热量会使任何侥幸形成的化学分子被加速分解。
由于水的存在造成这许多问题,致力于研究生命起源的物理化学家格拉哈姆·凯恩斯-史密斯(Graham Cairns-Smith)提出,粘土表面能促进一些所需的反应。
然而,在温暖的火山边的水池中进行的实验表明,粘土颗粒与氨基酸、DNA和磷酸基等形成生命的组分结合得太紧密,因此粘土会阻止这些物质之间发生必要的反应。31
如上所述,含有DNA、蛋白质和RNA的能繁殖的细胞绝不会在化学汤中偶然发源。所以,倡导生命自然发生的学者们又想象出另一种情况:刚开始时生命的需求比较简单,然后逐步进化到如我们今天所知的生命。
先出现蛋白质?
大多数研究集中在“先出现蛋白质”的起源方式上。根据这种途径,蛋白质首先出现,然后出现了DNA序列并能够产生所需的蛋白质,随后从DNA序列制造了合成蛋白质所必需的RNA。然而,除了获得一整套100%手性正确的氨基酸材料的难题,以及氨基酸聚合制造蛋白质链的难题之外,几乎没有蛋白质可以作为模板来复制自己。32此外,正如信息理论家休伯特·约克奇(Hubert Yockey)所指出的那样,一个基本难题是没有任何机制能够从蛋白质制造编码这个蛋白质的DNA序列。33
先出现RNA?
在20世纪80年代,研究者发现了一些能够催化某些化学反应的RNA分子,称之为“核酶”(核糖核酸酶)。这一发现令人振奋,导致很多人将精力花费在“RNA第一”或“RNA世界”的理论上。至少有一些酶可以根据RNA信息制造DNA信息,也就是说,如果你能得到RNA,就可以想象出获得DNA的途径。然而,能够从RNA序列来制造DNA拷贝的酶复合物非常复杂,这些酶复合物本身绝不可能通过自然过程而出现。在“RNA第一”的理论中还有许多看似无法克服的其他困难,凯恩斯-史密斯列举了其中的19个难题。34 此外,如上所述,DNA非常不稳定,但RNA的稳定性比DNA差得多。
人们提出了许多种生命起源可能发生的过程,这反而强化了一个结论:研究人员其实不知道生命是如何“自我创造”的。对于生命是如何从简单状态开始并逐步进化成一个真正的活细胞,没有人能提出可行的假说。新达尔文主义(即突变和自然选择)常被用来“跨越无望巅峰”,但是在一个能够存活且能自我繁殖的实体(即细胞,我之前在“细胞存活的最低要求是什么?”一节中已经列举了它的最低需求)出现之前,突变和自然选择从理论上就完全不起作用。
生命来自外太空?
弗朗西斯·克里克(Francis Crick)是DNA双螺旋结构的发现者之一,也是“生命源自外太空”的著名支持者。35 他提出外星人将生命送到地球,称为“定向泛种论(directed panspermia)”。这种想法的另一种形式,简称“泛种论(panspermia)”,就是生命在宇宙中的其他地方出现,并以微生物的形式,通过陨石或彗星来到地球,以这种方式向地球“播种”了生命。两种版本的泛种论都有效地将生命起源的难题推出了科学的范围以外。关于泛种论假说,唯一可测试的环节是微生物在冲向地球的陨石上或在陨石中的存活能力。这个问题已经过测试,结果对泛种论不利,微生物无法存活。36
人们进行外星文明探索(SETI)和太阳系外行星探索的原动力在很大程度上是为了发现生命在天外形成的证据。但即使将整个宇宙作为实验室也无法解决生命起源的问题:生命永远不会自发形成,如下所述。
生命起源的概率计算
对于化学物质演化成生命的概率,已经有许多人进行了尝试性的计算,但所有这些尝试都使用了简化的、让生命自发起源更有可能性(即概率> 0)的假设。
 数学家弗雷德·霍伊尔(Fred Hoyle)爵士以各种方式说明了生命自然形成的极端不可能性,甚至连一种有功能的聚合物分子,如蛋白质,也不可能自发形成。霍伊尔说,“现在想象有1050个盲人[编辑补充:如果他们肩并肩站立,整个地球系都容不下他们],每个人都转动着一个被打乱的魔方,试着想象他们同时让魔方各面都转到相同颜色的几率。如果上述目标能够达到的话,你才有机会通过随机排列的方式,获得生命所依赖的众多生物聚合物中的一个。除了生物聚合物之外,就连活细胞的操作程序(遗传信息)也要在地球上的原始汤中随机形成,如此概念简直是痴人说梦。生命必定是一种宇宙现象。“ 37
数学家弗雷德·霍伊尔(Fred Hoyle)爵士以各种方式说明了生命自然形成的极端不可能性,甚至连一种有功能的聚合物分子,如蛋白质,也不可能自发形成。霍伊尔说,“现在想象有1050个盲人[编辑补充:如果他们肩并肩站立,整个地球系都容不下他们],每个人都转动着一个被打乱的魔方,试着想象他们同时让魔方各面都转到相同颜色的几率。如果上述目标能够达到的话,你才有机会通过随机排列的方式,获得生命所依赖的众多生物聚合物中的一个。除了生物聚合物之外,就连活细胞的操作程序(遗传信息)也要在地球上的原始汤中随机形成,如此概念简直是痴人说梦。生命必定是一种宇宙现象。“ 37
实际上,我们可以计算出只获得一个长度为150个氨基酸的小蛋白质的概率,假设只存在(手性)正确的氨基酸,并假设它们将以正确的方式连接在一起(聚合)。有20种不同的氨基酸,150个氨基酸的可能排列的数量是20150 ,或者说得到正确的蛋白质的概率约为 10195分之一 。为了避免有人反对说:不是每个氨基酸都必须按照确切的顺序排列,这只是使用一种小蛋白质进行计算,并且只计算几百种生命必须蛋白质(许多蛋白质的长度大得多)中的一种,如果同时计算DNA序列自发形成的概率,会使这个问题严重地复杂化。实际上,有些蛋白质的序列即使发生了少量改变,也会失去全部功能。38
那时霍伊尔认为生命必须来自外太空。后来他意识到,在没有指导(没有智慧参与)的条件下,即使将整个宇宙作为实验室,生命也不可能在任何地方通过物理和化学的过程而形成:
“从无生命物质中形成生命的可能性是104,000分之一……它足以埋葬达尔文和整个进化论。没有原始汤,在这个星球上和任何其他星球上都没有,如果生命的起源不是随机的,那么生命肯定是有目的的智慧产物。“ 39
如果没有目的和智慧,那么104,000分之一这个数字会不会否定生命在宇宙中任何地方发生的可能性?我们可以这么说吗?
麻省理工学院研究员塞思·劳埃德(Seth Lloyd)计算了所谓的宇宙大爆炸(据称是在137亿年前发生的)之后的一切可能发生的事件(或“基本逻辑运算”)的总数量,结果是不超过10120。40 这就设定了理论上可能进行的实验数量的上限。此上限意味着概率为104,000分之一的事件将永远不会发生,即使是我们计算的一种150个氨基酸的小蛋白质在整个宇宙中也不可能自发形成。
然而,生物物理学家哈罗德·莫罗维茨(Harold Morowitz)41 得出的概率要低得多,为1010,000,000,000分之一。这是从含所有细菌基本零件的肉汤(例如,理论上可以通过加热含活细菌的培养液,杀死它们并将它们分解成基本成分而获得)自发组装而得到最简单细菌的机会。
莫罗维茨是无神论者,但他认为生命不是偶然因素的结果,而是出自可用之能量的某种属性,该属性驱动了能够使用能量的实体(即“生命”)的形成。这听起来很像地母盖亚的神话,因它为宇宙赋予了泛神论的神秘属性。
最近,无神论哲学家托马斯·内格尔(Thomas Nagel)提出了类似的旨在解释生命和思想之起源的说法。42
只要不用相信一个超自然的创造者,好像什么理论都可以提出来。
由于概率计算的困难,使用不同的假设,就会计算出不同的概率。如果我们使用了最有利于生命自然发生的假设进行计算,其结果仍然是极端的不可能,那么这就是一个比使用更现实的假设更有力的论证。更现实的假设虽然让唯物主义的说法更加不可能,但唯物主义者会质疑我们的某些假设。
然而,所有关于生命之化学起源的概率计算都会使用不切实际的、有利于生命化学起源的假设,否则概率就会为零。例如,莫罗维茨计算中使用的含有活细胞的所有成分的肉汤是不可能存在的,因为如上所述,这些化学成分会互相反应,使得它们不能用于制造活细胞所需的复杂聚合物。
著名信息理论家休伯特·约克奇(加州大学伯克利分校)认识到了这个问题:
“原始汤中生命随机起源在概率上是不可能的,就像永动机在概率上不可能一样。虽然本章计算的极小概率并不会令真正的信徒沮丧…… [然而]一个现实的人必会得出结论:生命并非偶然形成的。”43
请注意,在约克奇的计算中,他慷慨地假设生命原料可以在原始汤中获得。但是在他书的前一章中,约克奇证明原始汤本身就根本不存在,所以相信原始汤是一种“信仰”。他后来得出结论,“原始汤理论是卫道士们基于其意识形态的自我欺骗。”44
更多的坦白
请注意,约克奇并不是唯一一位在这个问题上明确发表言论的知名学者:
“如果任何人告诉你,他或她知道34亿年前地球上的生命是如何开始的,这个人不是一个傻瓜,就是一个骗子。没人知道。” ——斯图亚特·考夫曼(Stuart Kauffman)教授,生命起源研究者,加拿大卡尔加里大学(University of Calgary)。45
“……我们必须承认,关于任何生化系统或细胞系统的演变,目前尚没有详细的达尔文主义的解释,而只有各种一厢情愿的推测。” ——弗兰克林·哈罗德(Franklin M. Harold),科罗拉多州立大学生物化学与分子生物学荣誉教授。46
“关于从非生命到生命的进化途径,达尔文曾写道:‘目前生命起源只是垃圾思想,还不如想想物质的起源。’我们今天几乎处于同样的黑暗中”——亚利桑那州立大学超越科学基本概念中心主任保罗·戴维斯。47
“细胞的新颖性和复杂性远远超过当今世界上任何无生命的东西,以至于我们不知道它是如何实现的。” ——克斯纳(M.W. Kirschner, 哈佛大学医学院系统生物学系主任、教授)和格哈特(J.C. Gerhart, 加州大学研究生院教授)。48
“结论:生命起源的科学课题可以归结为寻找一个化学机制,能够实现从第一个自动催化的自我复制周期开始,最后演化到一个共同祖先。目前所有的理论都远远未能完成这项任务。虽然我们仍然不清楚这种机制,但我们现在已经理解了这个问题的广度。” 49
“进化论的最大缺口仍然是生命本身的起源……从这一堆分子[氨基酸和RNA]到即使是最原始的细胞,差距仍然是巨大的。”——克里斯·威尔斯(Chris Wills),加州大学生物学教授。50
即使是唯物论教条主义者理查德·道金斯(Richard Dawkins)也向本·斯坦因(Ben Stein,见电影纪录片Expelled)承认没有人知道生命是如何开始的:
道金斯:“我们知道生命起源中必定发生的一件事,就是出现第一个自我复制的分子。“
斯坦因:“那是怎么发生的?“
道金斯:“我告诉过你,我们不知道。”
斯坦因:“所以你不知道它是如何开始的?“
道金斯:“不知道,没人知道。” 51
“我们永远不会知道生命最初是如何出现的。 然而,对生命起源的研究是一个成熟、完善的科学研究领域。正如进化生物学的其他领域一样,对于最初生命形式的起源和本质问题的答案,只能算是探索和解释,而不是明确的和肯定的。” 52
结论
 若没有智慧,单凭物理过程和化学反应就不会产生生命。创造生命,即使是最简单的生命,所需的智慧都远远超过人类的智慧;人类仍然在摸索着试图完全理解最简单的生命形式是如何运作的。即使是最简单的细菌中也还有很多东西需要我们学习。事实上,随着我们学到的知识越来越多,生命起源的“问题”变得更加困难;答案不是越来越接近,而是越来越遥远。但真正的问题在于:生命的起源大声告诉我们有一个超级智慧的生命创造者,而这正是今天的世俗头脑所不能接受的。
若没有智慧,单凭物理过程和化学反应就不会产生生命。创造生命,即使是最简单的生命,所需的智慧都远远超过人类的智慧;人类仍然在摸索着试图完全理解最简单的生命形式是如何运作的。即使是最简单的细菌中也还有很多东西需要我们学习。事实上,随着我们学到的知识越来越多,生命起源的“问题”变得更加困难;答案不是越来越接近,而是越来越遥远。但真正的问题在于:生命的起源大声告诉我们有一个超级智慧的生命创造者,而这正是今天的世俗头脑所不能接受的。
若是科学能“证明”上帝的存在,生命的起源可以说是最好的证据。
相关文章
-
-
- Vintage Journal: The origin of life
- Evolutionist criticisms of the RNA World conjecture
- Origin of life: the polymerization problem
- Origin of life: the chirality problem
- Origin of life: instability of building blocks
- Physicist’s ‘breakthrough’ on the origin of life: can thermodynamics of heat dissipation explain chemical evolution?
- The Miller–Urey experiment revisited
- Reading ‘origin of life’ research
-
相关录像
Origin Of Life-Evolution Achilles Heels
参考与注释
-
- http://evolution.berkeley.edu/evosite/evo101/IIE2aOriginoflife.shtml (accessed 17 October 2013).
- Myers, P.Z., 15 misconceptions about evolution, 20 February 2008, scienceblogs.com; Matzke, N., What critics of neo-creationists get wrong: a reply to Gordy Slack, pandasthumb.org. 道金斯在他自称“证明了进化论“的著作《地球奇观》(The Greatest Show on Earth)中试图解决生命起源的问题。参见 Sarfati, J., The Greatest Hoax on Earth? ch. 13, 2010, Creation Book Publishers.
- Kerkut, G.A., Implications of Evolution, Pergamon, Oxford, UK, p. 157, 1960 (在线阅读ia600409.us.archive.org/23/items/implicationsofev00kerk/implicationsofev00kerk.pdf); creation.com/evolution-definition-kerkut.
- 糖具有含羰基的线性结构 – 参见图2. 核酸中出现的环状结构的糖在溶液中也占优势,但与线性结构处于平衡状态。当某些物质与醛基强烈反应时,会再生出更多的线性糖以代替反应消耗掉的那些线性糖,最终所有的糖分子将被消耗掉。
- Sarfati, J., World record enzymes, Journal of Creation 19(2):13–14, 2005; creation.com/world-record-enzymes-richard-wolfenden.
- Bergman, J., Why the Miller-Urey research argues against abiogenesis.
- Truman, R., What biology textbooks never told you about evolution.
- Sarfati, J., Origin of life: instability of building blocks.
- Chadwick, A.V., Abiogenic Origin of Life: A Theory in Crisis, 2005; origins.swau.edu/papers/life/chadwick/default.html.
- 作为一例,见Potassium ion channel, hydrated ionic radii, creation.com/ionic-error, 21 August 2010.
- Krogh, A. et al., Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes, Journal of Molecular Biology 305(3):567–580, 2001; dx.doi.org/10.1006/jmbi.2000.4315.
- 生殖支原体G-37中的转运蛋白; 见membranetransport.org/index.html, 2013年10月11日。
- 手性中的“右”和“左”是指氨基酸的(费歇尔投影)标准化结构图上显示的氨基(NH2) 处于右/左边的位置。
- Sarfati, J., Origin of life: the chirality problem; creation.com/origin-of-life-the-chirality-problem (updated 2010).
- Cairns-Smith, A.G., Evolutionist criticisms of the RNA World conjecture, from Genetic Takeover and the Origin of Life, 1982; creation.com/cairns-smith-detailed-criticisms-of-the-rna-world-hypothesis.
- Stanford researchers produce first complete computer model of an organism; news.stanford.edu, 19 July 2012.
- Sarfati, J., How simple can life be? https://creation.com/how-simple-can-life-be. 对人工合成细胞JCVI-syn3.0的研究显示, 473个基因是必需的,其中65个功能不明: C.A. Hutchison III et al., Design and synthesis of a minimal bacterial genome, Science 351:1414, March 25, 2016; doi: 10.1126/science.aad6253.
- Sarfati, J., Origin of life: the polymerization problem.
- Davies, P., The secret of life won’t be cooked up in a chemistry lab: Life’s origins may only be explained through a study of its unique management of information, The Guardian, Sunday 13 January 2013; guardian.co.uk/commentisfree/2013/jan/13/secret-life-unveiled-chemistry-lab. Return to text.
- Davies, P., Life force, New Scientist 163(2204):27–30, September 18, 1999.
- Popper, K.R., “Scientific reduction and the essential incompleteness of all science”; in Ayala, F. and Dobzhansky, T., (Eds.)., Studies in the Philosophy of Biology, University of California Press, Berkeley, p. 270, 1974.
- Smith, C., Lost in translation: The genetic information code points to an intelligent source, 6 May 2010; creation.com/genetic-code-intelligence.
- Freeland, S.J., et al., Early fixation of an optimal genetic code, Molecular Biology and Evolution 17(4):511–18, 2000; mbe.oxfordjournals.org/content/17/4/511.full.
- DNA有4个核苷酸“字母”(A、T、G、C),分子阅读机器一次读取3个字母(即一个“密码子”),如此就有4x4x4 = 64种可能的密码子排列。
- 通常有3个“终止”密码子作为蛋白质编码序列结束的标记,因此正常有61个密码子用于氨基酸编码。
- Novoa, E.M. and de Pouplana, L.R., Speeding with control: codon usage, tRNAs, and ribosomes, Trends in Genetics 28(11):574–581, November 2012; ww2.biol.sc.edu/~elygen/biol655/translation%20speed.pdf.
- Newly discovered DNA repair mechanism, Science News, sciencedaily.com, 5 October 2010. Return to text.
- Sarfati, J., New DNA repair enzyme discovered, 13 January 2010; creation.com/DNA-repair-enzyme. Tomas Lindahl, Paul Modrich, 和 Aziz Sancar 因发现三种DNA修复机制获2015年诺贝尔化学奖: Batten, D., DNA repair mechanisms ‘shout’ creation, Creation 38(2):56, April 2016.
- Cox, M.M., Keck, J.L. and Battista, J.R., Rising from the Ashes: DNA Repair in Deinococcus radiodurans, PLoS Genetics 6(1): e1000815, 2010; doi:10.1371/journal.pgen.1000815.
- Catchpoole, D., Life at the extremes, Creation 24(1):40–44, 2001; creation.com/extreme and Sarfati, J., Hydrothermal origin of life? Journal of Creation 13(2):5–6, 1999; creation.com/hydrothermal.
- Morelle, R., Darwin’s warm pond idea is tested, 13 Feb. 2006; news.bbc.co.uk/2/hi/science/nature/4702336.stm.
- 朊病毒有时被认为是能复制的蛋白质,但朊病毒是引起既有的蛋白质变形;它们不会通过使氨基酸按朊病毒自己的顺序排列来复制朊病毒(朊病毒被认为会导致“疯牛病”)。
- Yockey, H., Information Theory, Evolution and the Origin of Life, Cambridge University Press, 2005, pp. 118–119.
- Evolutionist criticisms of the RNA World conjecture; Quotable Quote by Cairns-Smith; creation.com/cairns-smith-detailed-criticisms-of-the-rna-world-hypothesis. See also, Mills, G.C. and Kenyon, D., The RNA World: A Critique, Origins & Design 17(1); arn.org/docs/odesign/od171/rnaworld171.htm.
- Bates, G., Designed by aliens? Creation 25(4):54–55, 2003; creation.com/aliens.
- Sarfati, J., Panspermia theory burned to a crisp: bacteria couldn’t survive on meteorite, 10 Oct 2008; creation.com/panspermia-theory-burned-to-a-crisp-bacteria-couldnt-survive-on-meteorite.
- Hoyle, Fred, The Big Bang in Astronomy, New Scientist 92:521–527, 1981.
- 例如,罗尔·杜鲁门(Royal Truman)研究了存在于真核生物中的泛素蛋白,结果表明,要保持其功能,蛋白质序列几乎不允许改变,因此这种蛋白质自然发生的可能性被排除了; 见Truman, R., The ubiquitin protein: chance or design? Journal of Creation 19(3):116–127, 2005; creation.com/the-ubiquitin-protein-chance-or-design。
- Sir Fred Hoyle, as quoted by Lee Elliot Major, “Big enough to bury Darwin”. Guardian (UK) education supplement, Thursday August 23, 2001; education.guardian.co.uk/higher/physicalscience/story/0,9836,541468,00.html.
- Lloyd, Seth, Computational capacity of the universe, Physics Review Letters 88:237901, 2002; http://arxiv.org/abs/quant-ph/0110141v1.
- Morowitz, H., Energy Flow in Biology, Academic Press, NY, 1968.
- Nagel, T., Mind and Cosmos: Why the Materialist Neo-Darwinian Conception of Nature Is Almost Certainly False, Oxford University Press, 2012.
- Yockey, H., Information Theory and Molecular Biology, Cambridge University Press, 1992, p. 257.
- Ibid. p. 336; see Quotable quote: Primeval soup—failed paradigm.
- Stuart Kauffman, At Home in the Universe: The Search for the Laws of Self Organization and Complexity, Oxford University Press, p. 31, 1995.
- Harold, F.M., The way of the cell: molecules, organisms and the order of life, Oxford Uni. Press, New York, p. 205, 2001.
- Davies, Paul, The Cosmos Might Be Mostly Devoid of Life: We still have no idea how easy it is for life to arise—and it may be incredibly difficult, Scientific American, 1 September 2016; http://www.scientificamerican.com/article/the-cosmos-might-be-mostly-devoid-of-life.
- Kirschner, M.W. and Gerhart, J.C., The plausibility of life: Resolving Darwin’s Dilemma, Yale University Press, New Haven and London, p. 256, 2005.
- Watchershauser, G., Origin of life: RNA world versus autocatalytic anabolism, The Prokaryotes, Vol. 1, 3rd edition, chapter 1.11, pp. 275–283, p. 282, 2006.
- 被引于Evolution’s final frontiers, New Scientist 201(2693):42, 2009.
- Expelled: no intelligence allowed, Premise Films, 2008.
- Lazcano, Antonio, Historical Development of Origins Research, Cold Spring Harbor Perspectives in Biology 2(11): a002089, November 2010; doi: 10.1101/cshperspect.a002089.
